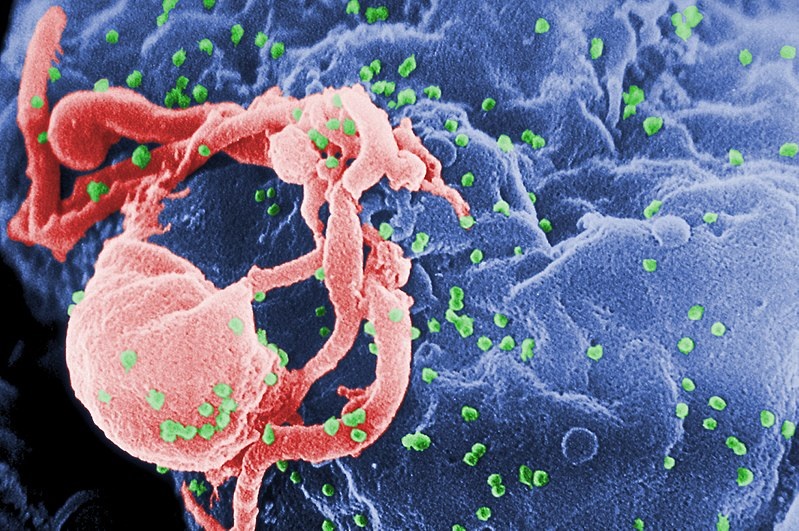
上図に培養リンパ球からの何千個ものHIV-1のビリオン(緑色)が出芽している走査型電子顕微鏡写真を載せます。赤色の細胞表面の複数の丸い隆起は、ビリオンの集合と出芽の部位を表しています。ビリオン (virion)とは、完全な形をしたウイルス粒子であり、増殖したHIVウイルスがビリオン(virion)となって、細胞外に放出されたウイルス (virus)の状態であり、完全な粒子構造を持ち、感染性を有するウイルス粒子がビリオンです。HIVはウイルスの分類上は、エンベロープを持つプラス鎖の一本鎖RNAウイルスであるレトロウイルス科レンチウイルス属のレトロウイルスです。一本鎖プラス鎖RNAウイルスはウイルスゲノムRNAがそのまま翻訳されウイルスタンパク質が作られますが、一本鎖マイナス鎖RNAウイルスではウイルスに由来するRNA合成酵素により相補的なRNAが合成されてのちウイルスタンパク質の発現が可能になります。 HIVはヒト免疫不全ウイルス(human immunodeficiency viruses、略してHIV)であり、遺伝子学的に、HIV-1 (Human Immunodeficiency Virus type1)と HIV-2 (Human Immunodeficiency Virus type2)の2種類のサブタイプに大別され、HIV-1は日本を含めた世界で最も流行している型です。HIVは後天性免疫不全症候群(AIDS)を引き起こし、その結果、生命を脅かす病原性が弱い弱毒菌が起こす日和見感染症やさらに癌を発生させることがあります。治療なしでは、HIV感染後の平均生存期間は、HIVサブタイプにもよりますが、9年から11年です。HIVサブタイプとは何でしょうか? HIV-1は変異を繰り返し起こしてさらに新たなる仲間を生み出し、その仲間の多くを占めるグループM(Main)と、限局した地域で認められるグループO(Outlier、訳として「中心からはずれた」)と、N(New)の3種に分類され、グループMはさらに9種のサブタイプ(A〜D,F〜H,J,K)に分類されます。このように、HIVの分類が複雑多岐になったのは、HIVが感染し増殖する過程で変異し、サブタイプ間等の遺伝子組換えが起こりやすいことが原因です。SARS-Cov-2(COVID-19)と似ていますね。ひょっとすればHIVの遺伝子を一部組み入れたかもしれませんね。アハハ!SARS-Cov-2(COVID-19)は感染を繰り返してイギリス型や南アフリカ型、インド型、ブラジル型の変異株が全世界にとめどなく広がっていますね。この変異は人類が消滅するまで続くでしょう。中国のように漢方の清肺排毒湯の煎じ薬を大量に服用すればSARS-Cov-2 を自分の免疫で治せるのに残念ですね。
ほとんどの場合、HIVは性感染症であり、性器からの接触感染や血液や尿道球腺液や精液や膣液を介して感染します。尿道球腺液とは、英語でpre-ejaculate fluidといい、性感のクライマックス前の性的刺激中に男性の尿道球腺によって生成される液体です。ejaculateというのは「射精する」という意味で、preは「前」ですから射精前の液体であり尿道に出すのでpre-ejaculate fluidは尿道球腺液と訳されるのです。この液体は尿道を出て、性的興奮の間に陰茎の先端に出ていくのがしばしば見られます。尿道球腺とは、カウパー腺とも言われ、前立腺の下にあるペア(一対)のエンドウ豆サイズの腺です。尿道を性感のクライマックスに向けて準備する透明な潤滑液です。AIDSに感染した母親から妊娠中や出産中に胎児や乳児が血液や膣液に曝露されたり、さらに母乳を介して、乳児へのHIVの非性的感染が発生することもしばしばあります。これらの体液内で、HIVは細胞に感染していない粒子としてや、感染した免疫細胞内にHIVウイルスとして存在しています。
HIVは、ヘルパーT細胞(CD4+T細胞)、マクロファージ、樹状細胞など、ヒトの免疫系の重要な細胞に感染するので免疫の働きが傷害を受け免疫が低下していくのです。特にHIVは次のようなメカニズムでCD4+T細胞 を減少させます。そのメカニズムの1つ目は感染したCD4+T細胞 をピロトーシス(pyroptosis)させることによって、2つ目は隣接する未感染の CD4+T細胞 をアポトーシスさせたり、3つ目は感染したCD4+T細胞 をウイルスが溶解感染(lytic infection)によって直接殺すことによって、4つ目は感染細胞を認識するCD8+T細胞 (CTL)により感染したCD4+T細胞を殺戮する4つのメカニズムがあります。CD4+T細胞の数があるレベル以下に減る時はCD4+T細胞であるヘルパーT細胞の免疫(細胞性免疫)が失われてしまうので日和見感染になりやすくなり、最後はAIDSの発症に繋がってしまうのです。
ピロトーシス(pyroptosis)は、細胞内ウイルス感染に最も頻繁に発生する炎症性の溶解性のプログラム細胞死です。細胞死が行われる細胞死の形態は3種類あり、アポトーシスとピロトーシスとネクローシスの3つです。アポトーシスとネクローシスの違いについてはまたの機会にしましょう。
HIVのサブタイプ
HIV種の比較
| 種 | 毒性 | 感染力 | 有病率 | 推定される起源 |
| HIV-1 | 高い | 高い | グローバル | 一般的なチンパンジー |
| HIV-2 | 下 | 低 | 西アフリカ | スーティーマンガベイ |
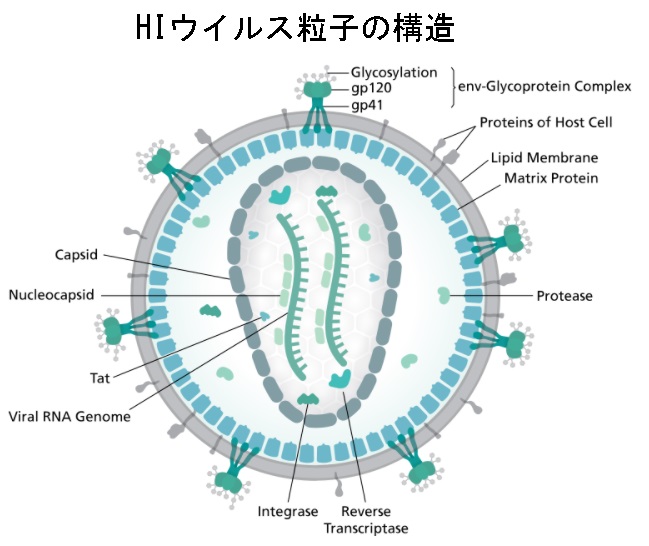
上のHIVは、レトロウイルス科のレンチウイルス属の一員です。多くの種がレンチウイルスに感染しており、レンチウイルスは潜伏期間の長い長期間の病気の原因となるのが特徴です。レンチウイルスは一本鎖のポジティブRNA(+鎖)でエンベロープ(Envelope、略してenv)をもった RNAウイルスです。env-Glycoprotein Complexというスパイクを利用して標的細胞に入るとすぐに、ウイルスのRNA ゲノム(Viral RNA Genome)はウイルスのRNAにコードされた酵素である逆転写酵素(Reverse Transcriptase)によって二本鎖DNAに変換(逆転写)され、ウイルス粒子(Virus Virion)内にウイルスゲノムとともに輸送されます。その結果、得られたウイルスDNAは細胞核に輸送され、ウイルスにコードされた酵素であるインテグラーゼ(Integrase)と宿主の補因子によって宿主細胞のDNAに組み込まれます。ひとたびDNAに組み込まれてしまうと、ウイルスは潜伏し、ウイルスとその宿主細胞が宿主細胞の免疫系により検出されることを不確定な時の間、回避することを可能にします。HIVウイルスは一次感染後、最大10年間、人体に潜伏し続ける可能性があります。この期間中、ウイルスは症状を引き起こしません。その代わりに、組み込まれたウイルスDNAは転写され、宿主細胞の持っている材料を利用して新しいRNAゲノムとウイルスタンパク質が生成されます。新しいRNAゲノムとウイルスタンパク質はパッケージ化(包装)され、複製サイクルを新たに開始する新しいウイルス粒子として細胞から放出されます。上図の中のTatの意味は何でしょうか? Tatは英語でTrans-Activator of Transcriptionの略で、訳すと転写の細胞間活性化因子となります。トランス活性化はTrans-Activatorとかトランス活性化因子とも呼ばれる内因性のウイルスタンパク質によって引き起こされます。 これらのタンパク質因子はトランスで(すなわち、分子間で)作用します。 HIVとHTLVは、ウイルス遺伝子発現を増強するトランス活性化因子をコードする多くのウイルスの2つの例です。Tatは、HIV-1のtat遺伝子によってコードされるタンパク質です。 Tatは、ウイルスの転写効率を大幅に高める調節タンパク質です。 このタンパク質は、サブタイプに応じて86〜101個のアミノ酸で構成されています。
HIVは他のレトロウイルスとは構造が異なります。ほぼ球形で、直径は約120nmで、赤血球の約60分の1です。これは、ウイルスタンパク質p24の2,000コピーで構成される円錐形のキャプシドで囲まれたウイルスの9つの遺伝子をコードするポジティブセンス一本鎖RNAを2本持っているのは上図でお分かりでしょう。一本鎖RNAは、ヌクレオカプシドタンパク質であるp7と逆転写酵素やプロテアーゼやリボヌクレアーゼやインテグラーゼのようなビリオンの発生に必要な酵素にしっかりと結合しています。ウイルスタンパク質p17から成り立っているマトリックス蛋白がビリオン粒子の保全を守るキャプシドを取り巻いています。次にマトリックス蛋白はウイルスエンベロープに囲まれています。これは新しく形成されたウイルス粒子が細胞から出芽したときに、ヒト宿主細胞の膜から取り込んだ脂質二重層で構成されています。ウイルスエンベロープは、宿主細胞からのタンパク質と比較的少数のHIVエンベロープタンパク質のコピーから得られたタンパクを含んでいます。このHIVエンベロープタンパク質はgp120(Glyco protein 120、略してgp120、訳して糖タンパク質120)と、3つのgp41分子(Glyco protein 41、略してgp41、訳して糖タンパク質41)から成り立っている幹から作られた蓋から成り立っています。gp120とgp41はその構造物をHIVのエンベロープに固定させています。エンベロープ蛋白はHIV env遺伝子によってコードされ、標的細胞に付着しウイルスのエンベロープを標的細胞を融合させるのです。その結果、ウイルスの内容物が細胞の中に放出され感染サイクルを開始されます。
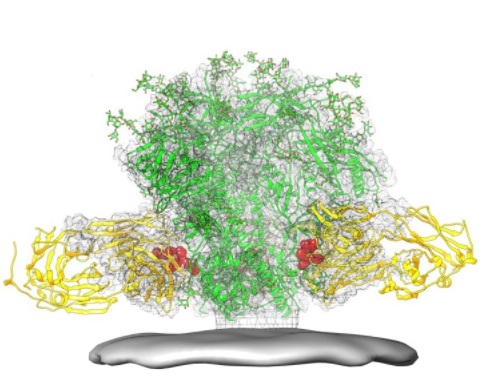
上図にHIVのスパイクタンパクの図を掲載します。 この図はHIVのスパイク蛋白に対するAIDSワクチンの働きの説明にもなります。融合ペプチドエピトープが赤で強調表示され、融合ペプチドに結合する広く中和する抗体(黄色)を含むHIVスパイクタンパク質(緑)も示されています。融合ペプチドとは何でしょう? HIVのエンベロープ蛋白はHIVが感染しようとしている人の標的細胞に付着しHIVウイルスのエンベロープを感染のための標的細胞のペプチドとを融合させるのです。その結果、ウイルスの内容物が細胞の中に放出され感染サイクルを開始されます。左上図の一番下の鉛色の板は標的細胞の細胞膜です。
ウイルス表面の唯一のウイルスタンパク質として、エンベロープのタンパク質はHIVワクチンの取り組みの主な標的です。三量体エンベロープスパイクの質量の半分以上は、N-結合型グリカンです。グリカンは、大きな炭水化物分子で 互いに化学的に結合した小さな糖分子を多数含んでいるのでpoly-saccharide(多糖類)とも呼ばれます。HIVウイルスの表面のエンベロープに含まれている多糖体であるグリカンは下にあるウイルスタンパク質を抗体による中和反応から保護するため、密度が高くなっています。これは、既知の最も密度の高いグリコシル化分子の1つであり、密度は十分に高いため、小胞体およびゴルジ装置での生合成中のグリカンの通常の成熟プロセスを妨げることができます。したがって、グリカンの大部分は、細胞表面に分泌または存在するヒト糖タンパク質には通常存在しない未成熟な「高マンノース」グリカンとしては完成されません。異常な処理と高密度は、これまでに同定されたほとんどすべての広く中和抗体(何ヶ月から何年も感染した患者のサブセットから)がこれらのエンベロープグリカンに結合するか、このようなエンベロープのグリカンに結合するか対処するように適応していることを示しています。中和抗体とは何でしょうか?
抗 体にはさまざまな働きがあります。抗体には異物(抗原)を分解する作用はありませんが、補体やマクロファージ、好中球などの貪食細胞を活性化して異物を排除します。つまり抗体は、異物を認識して結合する働きと、免疫を担う細胞を活性化させて異物を排除する働きがあります。抗体の働きを大きく分けると、異物(抗原)の中和作用、オプソニン化、細胞溶解、炎症の誘発の4つがあります。抗体の異物(抗原)に対する中和作用には2つあります。1つ目が特異的な抗体がウイルスのスパイクと結びつくとそのウイルスが細胞に感染できなくなります。2つ目はたとえ抗体と結びついたウイルスが細胞に感染してもそのウイルスが増殖することができなくなるのです。
HIVウイルススパイクの分子構造は、X線結晶学および極低温電子顕微鏡法によって決定されました。HIVウイルスのゲノムについても明らかになっており、HIV-1のRNAゲノムの少なくとも7つの特徴的な構造タンパクを作るのに必要なDNA配列には7つあり、a)LTR、b)TAR、c)RRE、d)PE、e)SLIP、f)CRS、g)INSの7つと、さらに他のタンパクがコードされている9つの遺伝子であります。それは1)gag、2)pol、3)env、4)tat、5)rev、 6)nef、7)vif、8)vpr、9)vpuであり、実は10番目の遺伝子tevが存在しています。10番目のtevは3)env、4)tat、5)revの遺伝子が融合し、19個のタンパク質をコードしている遺伝子です。これらの10個の遺伝子のうちの3つの遺伝子である1)gag、2)pol、3)envに、新しいウイルス粒子(ビリオン)の構造タンパク質を作成するために必要な情報が含まれています。ちなみに、DNA配列のa)のLTRは、英語でLong terminal repeat、略してLTRとなり、訳して「長い末端反復」であり、同じ配列が数百から数千回繰り返すDNA配列で、レトロトランスポゾンやレトロウイルスなどのRNAが逆転写されて生じるプロウイルスDNAの両端にみられます。 プロウイルスとは、宿主細胞のDNAに組み込まれてしまったHIVウイルスゲノムのことです。LTRはウイルスが自身の遺伝物質を宿主ゲノムに挿入するために用いられます。DNA配列のb)のTARは、英語でTrans-activation response elementで、訳してトランス活性化応答エレメントであり、26塩基対から成り立っています。TARはウイルスタンパク質と相互作用を通じて、転写活性化させる必須の役割を担っています。DNA配列のc)のRREは、英語でrev responsive elementで、Rev が結合し完全長のウイルスゲノム RNA が効率よく核から細胞質へ輸送されるのに必要です。Revは、5番目の遺伝子のrevによってコードされているタンパク質で、HIV-1タンパク質発現の調節に不可欠なトランス活性化タンパク質です。核局在化シグナルはrev遺伝子にコード化されており、Revタンパク質を核に局在化させることができ、スプライスされていない、または不完全にスプライスされたmRNAの輸送に関わっています。
次にHIVの9つの遺伝子の3番目のenvは、細胞プロテアーゼによって2つに切断されてgp120とgp41を形成するgp160と呼ばれるタンパク質をコードしています。残りの6つの遺伝子、tat、rev、nef、vif、vpr、vpu(またはHIV-2の場合はvpx)は、HIVが細胞に感染し、ウイルスの新しいコピーを生成する能力を制御するタンパク質の調節遺伝子です。特に4番目のtatという遺伝子によってコードされるTatタンパク質は、ウイルスの転写効率を大幅に高める調節タンパク質です。Tatは、英語でTrans-Activator of Transcriptionの略です。2つのtatタンパク質(p16およびp14)は、TARRNAエレメントに結合することによって作用するLTRプロモーターの転写トランス活性化因子です。 プロモーターとは何でしょう? 特定の遺伝子の転写を開始するDNA配列領域をプロモーターと言います。もっと詳しく言うとプロモーターの下流にあるDNAから単一のRNAの転写を始めさせるタンパクと結びつくDNAの配列であります。
レトロウイルス科のレンチウイルス属の種であるHIV-2は、HIV-1よりも病原性がはるかに弱いので西アフリカだけに見られます。感染性病原体の生存戦略は、その宿主を殺すことではなく、最終的には共生生物になることですが、弱い病原性を達成した後、時間が経つにつれて感染しやすい変異体が生まれてきます。
ヘルペスウイルスの特異性は、あらゆるウイルスの中でヘルペスは感染性がもっとも多様で強力であるにもかかわらず、病原性が極めて低いので、一度感染すると殺されることもなく、宿主が死ぬまで増殖を続ける極めて特異的な病原性ウイルスなのです。ヘルペスウイルスは宿主の免疫が弱い時には共生しているふりをし、免疫が強くなると免疫から回避してエピゾームの形でDNAを輪環状にして逃げ隠れし続ける天才的なウイルスといえます。人類の病気の原因として最後まで残る病原体は、まさにこのヘルペスウイルス8種類なのです。にもかかわらず、ヘルペスウイルスが無視され続けるのは宿主である人間を殺すことがないからです。
HIVウイルスのライフサイクルの図
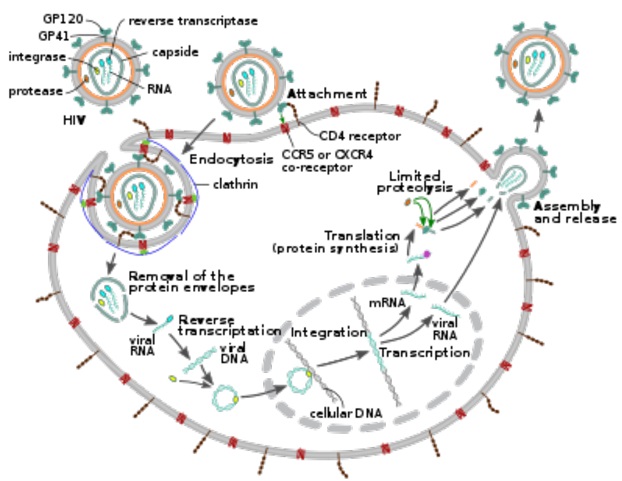
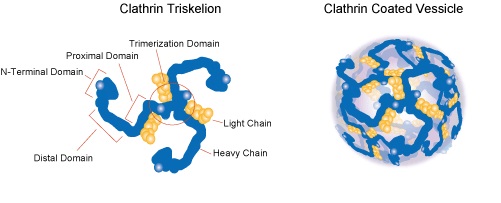
上図のブルーで示しているクラスリンについて説明しておきましょう。クラスリン (clathrin) は細胞外マトリクスの分子であるHIVウイルスが上図のようにエンドサイトーシスにより細胞に取り込まれる(感染する)際に形成される、エンドソーム外側を形作る骨格となるタンパク質であり、クラスリン分子は三脚巴構造 (tri-skelion) を取り、エンドソーム形成時は、複数のクラスリンが重合して格子を作り、サッカーボールの様な構造を形作ります。 三脚巴(さんきゃくともえ)とは、伝統的な文様のひとつであり、左にクラスリン分子の三脚巴構造 (tri-skelion)を示しておきましょう。三脚巴構造 (tri-skelion)は、三つの足を持っています。それぞれの足はブルーのクラスリン重鎖とイエローのクラスリン軽鎖から成り立っています。クラスリン重鎖は1675アミノ酸残基よりなり、分子量は約191kDaです。クラスリン軽鎖の分子量は約25-29kDaです。クラスリン重鎖は、ヘリックス-ループ-ヘリックス構造をとり、C末端側から、Trimerizationドメイン、proximal(近位)ドメイン、kneeドメイン、distal(遠位)ドメイン、ankleドメイン、およびterminal(終末)ドメインの6つのドメインより成り立っています。Trimerizationドメインは三脚巴構造の中心で、C末端側100残基程度で3つの足を相互につなぎ止めています。Terminalドメインはβ-プロペラ構造を取っており、被覆形成時には骨格の形成には関与せず、被覆の内側に配向しています。クラスリン軽鎖の生理的役割は、細胞分裂中期においては、有糸分裂紡錘体の動原体繊維(正確には微小管もしくは微小管結合タンパク質)と結合し、動原体繊維の配向(向き)を制御しています。さらに、ガン抑制因子であるp53タンパク質と結合し、p53の転写活性化能を制御することができるので癌を作りやすいのです。ク ラスリンを介したHIV-1のエンドサイトーシスによる感染が可能となるのです。
Mechanism of viral entry(ウイルス侵入の4段階のメカニズムの図)
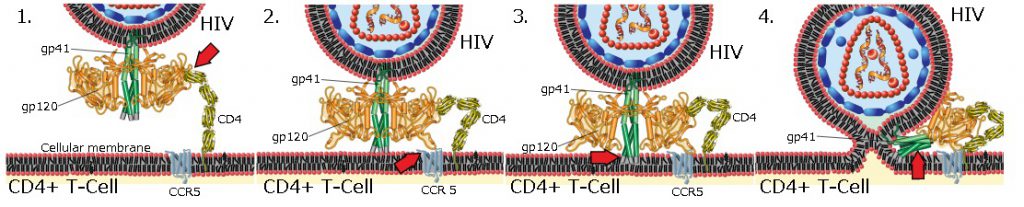
1)gp120とCD4+T細胞のCD4との最初の相互作用図 2)gp120のコンフォメーション変化により、CCR5との二次的な相互作用が可能になる図 3)gp41の遠位先端が細胞膜に挿入される図 4)gp41が大きな構造変化を起こす図 この時、半分に折りたたんでコイルドコイルを形成します。このプロセスは、ウイルス膜と細胞膜を引き寄せ、それらを融合させます。コイルドコイルはタンパク質の構造モチーフの1つで、2つから7つのαヘリックスがロープのように巻いた形をしています。コイルドコイル構造を持つタンパク質には、遺伝子発現を制御するものや転写因子など、重要な生物学的機能を持つものが多いのです。代表的なものにがん遺伝子由来のc-fosや、junや、筋肉中のトロポミオシンがあります。トロポミオシン(Tropomyosin)とは、アクチンの働きを調節する線維状のアクチン結合タンパク質であり、2本のαヘリックスからなるコイルドコイルの構造をとり、特に筋収縮を行う上で重要な働きをしています。トロポニン複合体が筋線維中のトロポミオシンに結合し、ミオシン結合を調節することで、筋収縮を調節しています。
HIVビリオンは、その表面の糖タンパク質が標的細胞の受容体に吸着し、続いてウイルスエンベロープが標的細胞膜と融合し、HIVキャプシドが細胞に放出されることにより、マクロファージおよびCD4+T細胞に侵入します。
HIVウイルスの細胞への侵入は、HIVウイルスエンベロープ上の三量体エンベロープ複合体(gp160スパイク)と、CD4および標的細胞表面上のケモカイン補助受容体(通常はCCR5またはCXCR4のいずれかです。他はCCR5とCXCR4の相互作用があります。)の両方の相互作用から始まります。gp160スパイクには、CD4受容体とケモカイン受容体の両方の結合ドメインが含まれています。
融合の最初のステップは、gp120のCD4結合ドメインのCD4への高親和性付着を含みます。gp120がCD4タンパク質と結合すると、エンベロープ複合体は構造変化を起こし、gp120のケモカイン受容体結合ドメインを露出させ、それらが標的ケモカイン受容体と相互作用できるようにします。これにより、より安定した2方面の付着が可能になり、N末端融合ペプチドgp41が細胞膜に浸透できるようになります。反復配列次に、gp41では、HR1とHR2が相互作用し、gp41の細胞外部分がヘアピンの形に崩壊します。このループ構造により、ウイルスと細胞膜が互いに接近し、膜の融合とそれに続くウイルスキャプシドの侵入が可能になります。
HIVが標的細胞に結合した後、HIV RNAと、逆転写酵素、インテグラーゼ、リボヌクレアーゼ、プロテアーゼなどのさまざまな酵素が細胞に注入されます。細胞の核への輸送中に、ウイルスの一本鎖RNAゲノムは二本鎖DNAに転写され、次に宿主染色体に組み込まれます。
HIVはこのCD4-CCR5経路で樹状細胞(DC)に感染します。樹状細胞(DC)は、性器感染中にウイルスが最初に遭遇する細胞の1つです。それらは現在、HIVウイルスがDCによって粘膜に捕捉されたときに、HIVが大好きなT細胞のCD4に取りつき、T細胞もHIVに感染してしまうのです。
HIVが細胞に感染した後、どのようにしてHIV-RNAがゲノムの二本鎖DNAへの逆転写を行うのでしょうか。
ウイルスキャプシドが細胞に入った直後に、逆転写酵素と呼ばれる酵素が、付着したウイルスタンパク質からポジティブセンス一本鎖RNAゲノムを遊離させ、それを相補DNA(cDNA)分子にコピーします。逆転写のプロセスは非常にエラーが発生しやすく、結果として生じる突然変異は薬剤耐性を引き起こしたり、ウイルスが体の免疫系を回避することを可能にします。逆転写酵素はまた、cDNAの合成中にウイルスRNAを分解するリボヌクレアーゼ活性、およびアンチセンスcDNAからセンスDNAを作成するDNA依存性DNAポリメラーゼ活性を持っています。cDNAとcDNAの相補配列が一緒になって二本鎖ウイルスDNAを形成し、それが感染T細胞核に輸送されます。ウイルスDNAの宿主細胞のゲノムへの組み込みは、インテグラーゼ(DNA統合酵素)と呼ばれる別のウイルス酵素によって実行されます。インテグラーゼ(DNA統合酵素)は、ウイルス由来のDNAが核内の宿主細胞DNAに組み込まれるのを触媒するウイルス酵素であり、活性化してウイルスタンパク質を産生できるプロウイルスを形成します。このように細胞の遺伝子に組み込まれたウイルスDNAは、HIV感染の潜伏期にあるため、休眠状態にあります。積極的にウイルスを産生するのには、細胞転写因子が必要であり、最も必要な転写因子はNF-κBであり、NF-κBはT細胞が活性化したときにアップレギュレート(上昇)します。NF-κBは積極的に感染と戦っているT細胞が活性化した時に、転写因子であるNF-κBも活性化して病原体と戦うべきなのに、逆にこのT細胞をHIVによって標的にされ、侵入され、その後免疫の王者であるT細胞が殺されてしまうという極めて皮肉な結果をもたらすのです。HIVは人間の免疫の遺伝子よりもはるかに賢いのでHIVは天才ですね。アハハ!
HIVウイルス複製中に、組み込まれたDNAプロウイルスはRNAに転写され、そのうちのいくつかはRNAスプライシングを受けて成熟したメッセンジャーRNA(mRNA)を生成します。これらのmRNAは核から細胞質に輸送され、そこで新しいウイルスの産生を促進する調節タンパク質TatとmRNAの輸送に関わるRevタンパク質に翻訳されます。新しく生成されたRevタンパク質が生成されると、Revタンパク質は核に移動し、そこでウイルスRNAの完全な長さではありますが、不必要な遺伝子を含んでいるスプライシングされていないコピーに結合し、それらが核を離れます。これらの完全長のRNAのいくつかは、ウイルスゲノムの新しいコピーとして機能しますが、他のRNAは、構造タンパク質GagおよびEnvを生成するために翻訳されるmRNAとして機能します。Gagタンパク質は、ウイルスRNAゲノムのコピーに結合して、新しいウイルス粒子にパッケージ化(部品の組み立てを)します。HIV-1とHIV-2は、RNAのパッケージ(部品の組み立て)が異なります。HIV-1は適切なRNAに結合します。HIV-2は、Gagタンパク質自体を作成するために使用されたmRNAに優先的に結合します。
2つのRNAゲノムが各HIV-1粒子にカプセル化されています。逆転写酵素によって触媒される感染と複製の際に、2つのRNAのゲノム間の組換えが起こります。組換えは、一本鎖プラス鎖RNAゲノムが逆転写されてDNAを形成するときに起こります。逆転写中に、複製初期のDNAはウイルスRNAの2つのコピーゲノム間で複数回組み換わってしまいます。この形式の組換えは、コピー選択として知られています。組換えイベントは、ゲノム全体で発生します。ゲノムごとに2〜20の組換えイベントが各複製サイクルで発生します。これらのイベントは、親から子孫のゲノムに送信される遺伝情報を迅速にシャッフル(入れ替える)する可能性があります。
逆転写酵素によるテンプレートスイッチングが一本鎖RNAゲノムの切断に対処するための修復プロセスとして機能しています。テンプレート(template、訳して鋳型や鋳型鎖)とは、DNAの複製やRNAの転写の際に配列が写し取られるもと(鋳型)となる鎖です。逆転写酵素(Reverse Transcriptase、略してRT)は、相同性に依存して1つのテンプレートから別のテンプレートに切り替えることができるのをテンプレートスイッチとか鎖スイッチングとかコピー選択組換えといいます。DNA合成中にテンプレートスイッチを作成する逆転写酵素の能力は、レトロウイルスのゲノム複製のモデル、ならびに組換えおよび癌遺伝子の形質導入が可能になります。
さらに、組換えはRNAゲノムの損傷の修復のためでもあり、逆転写酵素によるテンプレートスイッチング(鎖スイッチングとかコピー選択組換え)は、2つの損傷した一本鎖RNAゲノムコピーから損傷を受けていないゲノムDNAのコピーを生成します。HIVにおける上記のコピー選択組換え、つまり各HIV粒子が1つではなく2つの完全なRNAゲノムを含むようになったのは単に遺伝子を組み変えることによって多様なHIV遺伝子を作成するためだけではなく、RNAの損傷の修復プロセスのために進化論的に生じたのです。
HIV-1感染は宿主がHIVからの防衛のために、免疫のために慢性炎症と活性酸素種の産生が生じます。したがって、HIVゲノムは、二本鎖RNAではない一本鎖RNAの切断を含む酸化的損傷に対して脆弱です。HIVや一般的なウイルスが宿主に感染した時に宿主免疫の防御戦略の1つは、ウイルスゲノムに対して損傷を与えるために活性酸素種(reactive oxygen species、略してROA)を生成してウイルスに対抗します。したがって、ウイルスのテンプレートスイッチングによるコピー選択組換えはウイルスゲノム損傷の修復への適応なのです。全ての生命の根源はゲノムですから、ゲノムの損傷をHIVも恐れているのです。
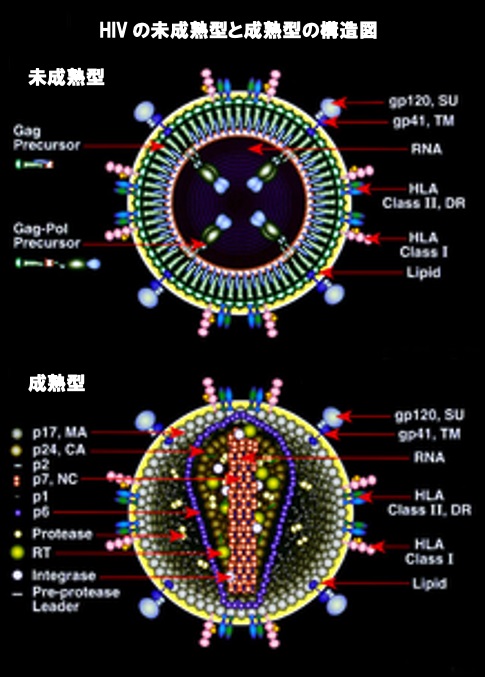
ウイルスのライフサイクルの最後のステップである新しいHIV-1ビリオンの組み立て(パッケージング)は、宿主細胞の原形質膜から始まります。Envポリタンパク質(gp160)は小胞体を通過し、ゴルジ装置に輸送され、そこでタンパクを活性化するフューリン酵素によって切断され、2つのHIVエンベロープ糖タンパク質であるgp41とgp120が生成されます。gp41とgp120は宿主細胞の原形質膜に輸送され、そこでgp41がgp120を感染細胞の膜に固定します。タンパク質が集まって出来たタンパク複合体(polyprotein、読みはポリプロテイン)であるGag(p55)およびGag-Pol polyprotein(p160)は、形成中のビリオンが宿主細胞から発芽し始めると、HIVゲノムRNAとともに原形質膜の内面にも結合します。Gag-Pol polyprotein(ギャグポリプロテイン)は、タンパク複合体なので実際のマトリックス、キャプシドおよびヌクレオキャプシドタンパク質に別々に切断される必要があるため、芽生えたビリオンはまだ未成熟です。この切断は、パッケージ化(組み立て)されたウイルスプロテアーゼによって媒介され、プロテアーゼ阻害剤の抗レトロウイルス薬によって阻害することができます。次に、さまざまな構造コンポーネントが集まって、成熟したHIVビリオンを生成します。成熟したビリオンだけが別の細胞に直接感染することができます。これを細胞から細胞へのウイルスの拡散といいます。
ビリオンによる細胞への感染は、感染T細胞で完成されたHIVビリオン(ウイルス粒子)は感染したT細胞から発芽し、血液またはリンパ液や細胞外液に入り、偶然に出会った別のT細胞に感染することもできます。これを無細胞拡散と呼びます。ヘルペスウイルスも細胞から細胞へと感染する経路と、まず増殖したヘルペスウイルスビリオンが血液やリンパ液や細胞外液に入って最後は別の細胞に感染する2つの経路があります。
HIVは、遺伝的多様性が非常に高いという点で多くのウイルスとは異なります。このHIVの非常に高い遺伝的多様性の理由は何でしょうか? まず1つ目は複製の一回のサイクルごとにヌクレオチド塩基あたり約3 × 10-5(10万分の3)の高い変異率があり、2つ目が逆転写酵素の遺伝子組換え特性があり、3つ目が1個のHIVが毎日約1010(100億個)のビリオンを生成するというとんでもないスピードの速い複製サイクルのためなのです。
この複雑なシナリオは、1人の感染した患者に1日の間にHIVの多くの変異亜種を生成することにつながります。今流行しているSARS-Cov-2も2週間で1回は変異しています。HIVもこのような多様性は、単一の細胞が2つ以上の異なるHIV株に同時に感染した場合にさらに悪化します。2つの変異株のHIVが同時に感染すると、子孫ビリオンのゲノムは、二つの異なる株からの違ったRNA鎖で構成されることもあります。このハイブリッドビリオンは新しい細胞に感染し、そこで複製されます。これが起こると、逆転写酵素は、2つの異なるRNAテンプレート(鋳型)間を行き来することによって、2つの親のゲノムの間の組換えである新しく合成されたレトロウイルスのDNA配列を生成してしまいます。この組換えは、様々のサブタイプ間で発生する場合に最も明らかとなるのです。
