抗体のクラススイッチと体細胞超突然変異が起こらないからです。IgMがどのようにしてIgGにクラススイッチするのか?体細胞超突然変異はどのようにして起こるのか?
「mRNAワクチンは絶対にできない。絶対に失敗する。 」も一緒にご一読ください。
SARS-CoV-2に対する獲得免疫の防御は3つの段階が必要です。1つ目は、コロナウイルスを認識できること。2つ目はコロナウイルスに合う武器(抗体)を産生すること。3つ目は、これらの武器をコロナウイルスの感染部位(肺)に運ぶことであります。獲得免疫の認識段階は二次リンパ器官で行います。二次リンパ器官には、リンパ節と脾臓と粘膜関連リンパ節(mucosa-associated lymphoid tissue、略してMALT)の3つがあります。一次リンパ器官は骨髄と胸腺です。



上 に二次リンパ器官の1つであるリンパ節の絵を掲載しておきます。この3つの二次リンパ器官の特徴は、リンパ濾胞(Lymphoid Follicle)を持っていることです。このリンパ濾胞(Lymphoid Follicle)には、一次リンパ濾胞と二次リンパ濾胞の2つがあります。まずリンパ節が活性化されない時には、一次リンパ濾胞として数多くのリンパ濾胞が左図のように存在しています。全てのリンパ濾胞には、濾胞樹枝状細胞(Follicular dendritic cell、略してFDC)が非常に多いのですが、最も多いのはBリンパ球なのです。従ってリンパ濾胞というのはB細胞という海の中に、濾胞樹枝状細胞(Follicular dendritic cell、略してFDC)の集団が島のようにあちこちに散在しているとイメージしてください。濾胞樹枝状細胞(Follicular dendritic cell、略してFDC)は、抗原を提示する樹枝状細胞(dendritic cell、略してDC)と同じくヒトデに似た格好をしていますが、2つの点で濾胞樹枝状細胞(Follicular dendriti ccell、略してFDC)と樹枝状細胞(dendritic cell、略してDC)は異なっています。
1つ目の違いは出生場所が異なり樹枝状細胞(dendritic cell、略してDC)は、白血球の1種類で骨髄で産生されます。その後、骨髄から血管を通って組織に出てコロナウイルスなどを見つける斥候の役目を果たします。一方、濾胞樹状細胞(FDC)は、受精後、妊娠中期までには二次リンパ器官であるリンパ節などに既に住みついています。
2つ目のDCとFDCの違いは、機能が異なっています。樹枝状細胞(dendritic cell、略してDC)の役割は、組織やリンパ節においてT細胞に抗原提示をする抗原提示細胞ですが、一方、濾胞樹枝状細胞(Follicular dendritic cell、略してFDC)の役割は、リンパ濾胞でB細胞に抗原を提示する抗原提示細胞なのです。
FDCがなぜB細胞に抗原を提示するのか不思議に思いませんか?皆さんは抗原提示細胞であるDCはMHCⅡに抗原ペプチドに乗せてヘルパーTリンパ球を活性化してIL-2、TNF、IFN-γなどのサイトカインを作ってナイーブTリンパ球をTh1リンパ球に変え、同時に病原体を殺しの抗体であるIgGを作らせるだけがBリンパ球の仕事だけではないかと考えておられるでしょう。
ところが元来、B細胞はヘルパーT細胞の手助けなしで、タンパク、炭水化物、脂質のみならず、あらゆる有機化学物質をBリンパ球の受容体で認識し、IgMを作り出すことができるのです。FDCが抗原をリンパ節のリンパ濾胞でBリンパ球に直接提示するのではなくて、FDCは提示した抗原をB細胞に捕まえさせ取り込ませる特別な仕事をしているのです。しかもFDCはDCと違ってMHCⅡを持っていないので、ペプチド抗原をMHCⅡと結びつけてヘルパーT細胞に提示することができないので並みのDCではなく、まさに濾胞でしか仕事ができない特殊なDCがFDCですから濾胞樹枝状細胞(Follicular dendritic cell、略してFDC)という名前が付けられたのです。ところがB cellはAPCの1つであるのでFDCと違ってMHCⅡ分子を持っており、かつCo-stimulator(補助刺激分子)であるのでリンパ節ではBリンパ球がヘルパーT細胞に対して抗原提示細胞(APC)になっているのです。
ところがあちこちですでに述べたようにヘルパーT細胞の手助けを借りないB細胞はIgMしか抗体を作れないのです。もちろん、IgG抗体のみならず、体細胞超変異も起こることもないのです。この体細胞というのはBリンパ球のことですよ。しかももっと正しくいえばB細胞のFabを決める遺伝子の塩基の並びが突然変異を起こすということです。B細胞のFabの突然変異が遺伝子に生じなければ、このB細胞はメモリーB細胞になることができないのです。ところがリンパ節のリンパ濾胞に住んでいる濾胞ヘルパーTリンパ球(Tfh)という特別なリンパ球が存在し、そのTfhの手助けを借りて上で述べたヘルパーT細胞に依存したB細胞でもできなかった仕事、つまり抗体のクラススイッチと体細胞超変異を起こさせることができるのです。
どのようにしてリンパ節での複雑極まりない免疫細胞の相互作用によってBリンパ球が最初に述べた3つの仕事をやり遂げるかをもう一度順にまとめましょう。この3つの仕事とは、1つ目は、コロナウイルスを完璧に認識でき、2つ目はコロナウイルスに対する完璧な武器であるIgG抗体を産生すること。3つ目は、これらの武器をコロナウイルスの感染部位(肺)に運ぶという3つの仕事です。この3つの仕事は必ずしも順番に起こる訳ではないことも理解して下さい。免疫学は医学の中で最も難しい分野であることは既に述べました。免疫学のハイライトは高等免疫(適応免疫)であり、この適応免疫がBリンパ球の3つの仕事を完成させる場所はリンパ節しかいないのです。

さてFDCの話に戻りましょう。FDCの仕事は何でしょうか?感染の初期に自然免疫の補体タンパクが新型コロナウイルスであるSARS-CoV-2と結びつくと、自然免疫の補体でオプソニン化された新型コロナウイルスであるSARS-CoV-2は二次リンパ器官であるリンパ節にリンパ液や血液によって運ばれます。二次リンパ器官(リンパ節)に住み着いているFDCはもちろん樹枝状細胞(dendritic cell、略してDC)の仲間の1つですから、補体と結びつくレセプターを持っているのでFDCは補体によってオプソニン化された抗原を奪い取り、保持し続けることが出来ます。実は保持するだけではなくて、それを取り込んで抗原タンパクをペプチドにできますがFDCは先ほど書いたようにMHCⅡはないので、そのペプチドをMHCⅡに乗せてBリンパ球に提示することはできないので、FDCはBリンパ球にオプソニン化されたSARS-CoV-2ウイルスのペプチドを提示すると、B細胞がペプチドをFDCから取り上げ、かつ取り込み、上の絵図のTh cellと結びつくとThはTfhという濾胞ヘルパーT細胞となり、このTfhと協力して最後はB細胞にクラススイッチと体細胞超変異を起こさせるのです。非常に大事なTfhについては後で詳しく説明します。
つまり、B細胞が作る抗体の種類を決めるFcの遺伝子の組み変えを行わせ(体細胞超変異)、IgMからIgGの遺伝子に変換して(抗体のクラススイッチ)、新しいIgGという抗体を作り替えさせるのです。
抗体のクラススイッチとは、IgMをIgGに変えることであり、さらにB細胞が作る抗体のFabの遺伝子を組み変えさせて、コロナウイルスとさらに強く結びつくFabの遺伝子を新たに作り替えることを体細胞超突然変異(Somatic super-mutationとかSomatic hyper-mutation)を起こさせるのです。体細胞突然変異(Somatic hyper-mutation、略してSHM)とは胚中心(germinal center)において、B細胞が高頻度にSHMを繰り返すことによって、抗体のコロナウイルス抗原とのアフィニティ(親和性の強さ)、アビディティー(結合力の強さ)が増強されていくプロセスを、アフィニティマチュレーションと呼びます。
アビディティーとアフィニティーの違いは何でしょうか?抗原抗体反応では、抗原上のエピトープ(抗体が認識する抗原の一部分)と抗体上のパラトープ(抗原結合部位であり抗原のエピトープを認識して結合する抗体の一部)が結合しています。抗体の可変領域のアミノ酸は約 110 アミノ酸から成り、軽鎖の可変領域と共に、抗原結合部位を構成します。そのアミノ酸配列とタンパク質の構造は抗体によって異なります。アフィニティー(Affinity)とは一価(一種類)のエピトープと一価(一種類)のパラトープとの結合力です。抗原が持つ一価のエピトープは数個のアミノ酸から出来ており、それに対応する抗体の一価のパラトープも数個のアミノ酸から成り立っています。実は抗原が持っているエピトープは一種類ではないのです。同じようにパラトープも一種類ではないのです。したがって抗原と抗体が1対1で対応して結合するアフィニティーを超えた抗原と抗体全体の総和的な結合力の概念が生まれたのです。その概念がアビディティーであります。アビディティーの方がアフィニティーよりも結合力がはるかに強力なのです。
アビディティー(Avidity)とは抗原と抗体の結合力の総和です。SHM(Somatic-hypermutation)による変異で生じた多様なB細胞群のうち、コロナウイルス抗原に曝露されることによりコロナウイルス抗原に対する高親和性抗体(Hyper-Affinity)を発現するB細胞がさらに増え維持され、他の低親和性抗体発現B細胞は排除されてしまうので、その結果、強い結合力を持ったB細胞だけが残っていくことをクローンセレクションといいます。しかし、SARS-Cov-2はいくつかのウイルスの遺伝子を組み込んだために変異が極めて起こりやすく、何百という変異種が出現しているのでクローンセレクションは起こりにくいので、ますますSARS-Cov-2は世界中を人間を苦しめることになるでしょう。
親和性とは何でしょうか? 親和性とは、1)エピトープに対する抗体の結びつきの強さ、2)抗原と抗体が結合しているエピトープの数、3)相互作用する部位の立体配置の適合性という主に3つの要因によって決められ、これらが、特異的な抗体がピッタリ合う抗原の持つエピトープに結合する可能性を高め、同じB細胞のクローンセレクションが生じるのです。
抗原がB細胞に取り込まれるまでは、FDC(Follicular dendritic cells、訳して樹状細胞)は人体に感染したコロナウイルスが増殖している肺組織で、補体と結合した新型コロナウイルスの持つ様々な種類の抗原でFDC(Follicular dendritic cells)の細胞膜はコロナウイルスの抗原だらけとなっています。一方、B細胞のBCR(B cell receptor)はウイルス抗原の上にある極めて小さいエピトープと結びついています。さらに大量のコロナウイルス抗原を捕まえて、それらを密に集めることによってFDCは、そのコロナウイルス抗原をB細胞に見せるときに、その抗原を認識するB細胞の受容体(BCR)が橋げたとなり、連続して抗原(エピトープ)に結びついて橋脚が繋がっているように見えるので、架橋する(橋を架ける)という表現を私は用います。このような架橋によってBCRが一カ所に集中して集まると抗原が大量に結びついたという情報が核に伝わりやすくなり、クラススイッチや超変異が起こしやすくなるのです。
いずれにしろ、このような架橋が見られるのは、3つの場合しかありません。1つは肥満細胞や好酸球や好塩基球が保有するIgEレセプターにIgE抗体が結びつく時に、そのIgE抗体のFabにアレルゲンのエピトープに結びつく場合と、2つ目はB細胞のレセプターである膜型IgM抗体のFab部分にオプソニン化された抗原のエピトープが結びつく場合と、3つ目は、B細胞の補体のレセプターの先端に補体でオプソニン化された抗原のエピトープが結びつく場合です。
元来、架橋という医学用語は、英語でcross linkといい、マスト細胞や好酸球や好塩基球上のIgEにアレルゲンが吸着し、多くのIgEにアレルゲンの橋がかけられた様に結びつけられると、この架橋による刺激によって脱顆粒が起こり、ヒスタミンが放出される意味で用いられました。
下の2つの図を見て下さい。上の絵図は、B細胞がヘルパーT細胞の助けを借りずに、初めて出会ったコロナウイルス抗原(Antigen)のcognate antigenのエピトープに対して膜型IgM抗体と結びついている絵図です。BCR(B cell receptor)は、訳してB細胞受容体ですが、膜型IgMそのものです。下の絵図はウイルス抗原(Antigen)のエピトープが、BCR以外に補体によってオプソニン化されたウイルス抗原を、もう1つの補体レセプターで認識しIgMを効率良く作れる状態を示しています。つまり敵であるコロナウイルス抗原をBCRと補体のレセプターの2つのレセプターで認識することができるので、この2つから伝えられたシグナル情報がB細胞の核に伝える力が2倍どころか何十倍も強くなり、B細胞は素早くIgM抗体を作れるようになるのです。この補体のレセプターをCo-receptor(補助受容体)といいます。しかしながら、2つのレセプターの刺激を得ても、あくまでもヘルパーT細胞があって初めて抗体のクラススイッチとさらに体細胞超変異(SHM)があってこそ強力な中和作用や親和性をもつIgGが作られるのです。しかもこの申し分のない最高の抗体であるIgGが作られる領域はまさにリンパ節の胚中心(germinal center)なのです。


さらにリンパ節の胚中心(germinal center)でこそ記憶B細胞、記憶T細胞、記憶キラーT細胞が生まれ、コロナウイルスに対するワクチンの効果が維持されるのですが、残念ながらコロナのスパイクタンパクの遺伝子情報を用いて作ろうとしているmRNAワクチンはこのような複雑極まりない機序を乗り越えてできるかについてはファイザーもアストロゼネカもバイオテックもモデルナも一言も言及していません。上の2枚の絵図とも一番上の直線が橋であり、Antigen(抗原)という一本の線が引かれていますね。この一本の橋の線が抗原であり、その橋にぶらさがっているのが抗原のエピトープであります。コロナウイルスのスパイク抗原(Spike Antigen)を一本の線の橋とみなして下さい。もちろんFDC細胞の手助けはまったくないので、コロナウイルスに対するIgM抗体だけが作れる可能性を示しているだけです。
世界中の全ての製薬メーカーがこのIgM抗体をIgG抗体であるかのような喧伝をやっていますが、たしかにIgM抗体が作られるのは確かです。抗体ができるという新聞記事は、あくまでもIgM抗体であるので、消えてしまうことがあることは当然なことなのです。しかしIgG抗体が決して作れないことや永久免疫が永久にはできないこと、つまり本当のワクチンができないことについては一言も触れいていないのは残念ですね。
何回も書きますが、ワクチンができるウイルスは、一回感染すると二度と同じ病気にならないという証拠が必要なのです。ところが今回のコロナウイルスは、一度かかっても再び感染することが世界中で報道されていますね。皮肉を言えば、ワクチンができるウイルスに感染することが、最高のワクチンになるのですがね。アッハッハ!しかし危険がありますね。
上の2つ絵図について追加説明を加えておきましょう。一本の線でAntigenと示されている抗原が、コロナウイルスの抗原(Virus Antigen)であり、このAntigenの線に数多くの薄赤の丸印がつけられているのがコロナウイルスの抗原のエピトープであり、エピトープ(epitope)にBCR(B細胞受容体)との結合に加えて、さらにB細胞のオレンジ色の棒状の補体レセプターが抗原と結合して2つの刺激によってB細胞が活性化され、その情報が核に伝えられ、膜IgM抗体(BCR)が分泌IgMを作るようになるのです。このIgM抗体の仕事は五量体になって補体を活性化することが1つで、かつ分泌抗体になったIgM抗体が血中に流れて、上手くいけばコロナウイルスと出会ってコロナのエピトープと結びつけば、感染の初期に中和作用としての働きが発揮されて、呼吸器感染症としての新型コロナウイルス(SRAS-CoV-2)が、呼吸器粘膜の細胞に感染しないようにすることもあり得ますが、あくまでもこのIgM抗体はmRNAワクチンが望む必要に応じて記憶免疫が作る永久抗体ではないのです。しかしワクチンとしては一時的には効果があるように見えますが、フェイクワクチンですね、アッハッハ!
すでに説明したように上図のエピトープ(epitope)というのは、コロナウイルス抗原の最小単位であり、抗体は新型コロナウイルスであるSARS-CoV-2などと結合する時、抗原の比較的小さな一部分のみを認識して、そのエピトープと結合する抗体の部分がパラトープ(Paratope)なのです。従って抗原と結びつくという表現は本来は正確ではなく間違いで、実は抗原の微小な一部であるエピトープとBCR(B cell receptor)の一部であるパラトープと結びつくのです。エピトープという言葉が難しいので簡単で大雑把な抗原という言葉でエピトープの代わりに用いるようになったのです。医学で一番難しいのは命を守る奥深すぎる免疫学なのです。私は、自分の若年性ヘルペス性脳炎という病気が進行しないようにしてくれた免疫を完全に理解すべく、楽しみながらこのように免疫学を勉強し、その成果を皆さんにAPCと同じように提示しているのです。アハハ!
ちなみにB細胞が補体のレセプターを持っているのは、B細胞自身がAPC(抗原提示細胞)であるので、樹状細胞と同じく補体のレセプターを持っているのは当然のことなのです。この補体のレセプターは、先に示したように、Co-receptor(コ-レセプターと読み、補助受容体)と言います。Co-receptorを共受容体と訳している医学書もありますが間違いです。何故ならば、補体のレセプターはあくまでもBリンパ球のBCRの働きを補助するので補助受容体と訳すべきなのです。
Co-stimulationという英語がありますが、これも共刺激と訳すのも間違いであり、正しくは補助刺激と訳すべきなのです。何故ならばヘルパーT細胞のCD40LがAPCであるB細胞のCD40と結合してB細胞を活性化する働きを補助的に手助けしているのでCo-stimulationは補助刺激と訳すべきなのです。
架橋のもう1つの例としては、アレルギーを起こす細胞である肥満細胞や好酸球や好塩基球にアレルギー抗体であるIgEが、結びついて、さらに大量のアレルゲンがIgEにひっついたというシグナル(信号)が核に伝わります。するとかゆみの原因となるヒスタミンが大量に肥満細胞から放出されてアレルゲンを体外から除去しようとするのです。
この時に細胞の膜に埋められているレセプターというのは実は膜の中で自由に移動が可能となり一カ所に集中するのです。このように受容体が集まることがクラスター(cluster)と言い、日本語で受容体の群れと訳します。レセプターを持っているすべての細胞はたくさんの情報を核に伝えるためにクラスターが起こっているのです。たとえばB細胞であるレセプターの膜抗体の細胞膜に埋めこまれているFc部分が近寄ることを膜抗体であるIgM受容体のクラスターといいます。一方、IgM膜抗体の細胞外に出て抗原と連続的に結びつく橋のように見えるのをクロスリンキング(cross linking)といい、架橋と訳すのです。BCRは実際には言うまでもなく連結しているわけではないのですが。したがってクロスリンキングとクラスターは同じ事と言えます。架橋という言葉は、化学反応において主に高分子化学においてポリマー同士を連結することです。免疫学における架橋という意味は、抗原に抗体が連続的に引っ付いているだけで、抗体自身が連結しているわけではないのです。
それではBCR(B cell receptor)やTCR(T cell receptor)は何のためにクラスターやcross linkingが免疫が進化して生まれたのでしょうか?
BCRのクロスリンキングとクラスターを取り上げて説明しましょう。
活性化シグナルを生み出すためには多くのBCRがB細胞の表面に密接に集まらなければならない必要があったからです。BCRがクラスター(群がる)するときに同時にクロスリンキング(連結)が生じているのです。抗原というのは、一連のアミノ酸の何回も繰り返されるタンパクから成り立っています。BCRは何も漠然と抗原と結合しているのではなくて、このように何回も繰り返される特異的なアミノ酸であるエピトープを認識しているのです。実際、抗原である細菌やウイルスや寄生虫の表面はいくつかの異なったタンパクのコピーから成り立っているのです。B cellのレセプターがこういうタンパクにあるエピトープを認識するとたくさんのBCRがクラスターするのです。BCRのクロスリンキング(架橋)はB cellの活性にとっては絶対に必要であるのです。何故でしょうか?さらに詳しく説明しましょう。今までは細胞膜の表に出ているY字型の抗体の頂上の2つの手の話ばかりでしたが、細胞の中に突き出ている尻尾がどのようになっているかについての話がまだでした。尻尾のタンパクはIgαとIgβというタンパクから成り立っています。抗原が両手のタンパクのH鎖とL鎖と引っ付いたという情報を細胞の中の核に伝えるために、このIgαとIgβは細胞の中にあるシグナルタンパク(信号を伝えるタンパク)と相互作用する必要があります。十分な相互作用が一カ所で集中したときに初めて酵素的な連鎖反応が開始され、細胞の核に信号を送ることができるのです。この信号伝達を送ることができるのは細胞の内のたくさんのIgαとIgβという分子が集まらなければならないのです。これがまさにB cell receptorのクロスリンキング(架橋)が行っている仕事なのです。つまりBCRのクラスター(集結)は充分なIgα分子とIgβ分子を密集させて、BCRに抗原が引っ付いたというシグナルを送る酵素的な鎖のような連続した反応を開始させることができるのです。
このように細胞内にあるレセプターの尻尾の多数のIgαとIgβが移動し密集するクラスター(集団)になると同時に細胞の外に出ているレセプターのYの両手が架橋すると、単独のレセプターが核に抗原と結びついたという情報を核に伝える力が100倍以上も強力になるのです。それでは難しすぎますがどのようにしてB細胞受容体がどのようにしてシグナル伝達を行うかを少し勉強しておきましょう。

上にB細胞受容体シグナル伝達経路の概略図を私自身のために掲載し、簡単な説明を加えておきます。
B細胞抗原受容体(Bcellantigenreceptor、BCR)は、膜結合型IgM免疫グロブリン(membraneimmunoglobulin、略してmIg)分子と、それに会合したIgα(CD79a)とIgβ(CD79b)ヘテロ二量体から成り立ちます。会合とは、同種の分子が分子間力によって2個以上結合し,一つの分子(単位)のように行動する現象であり、このような分子単位を会合体と呼びます。単位を構成する分子数によって二量体,三量体などと呼び、IgαとIgβは二量体になっているのです。mIgサブユニットは抗原に結合して受容体の凝集を起こし、一方でα/βサブユニットは細胞内へ向けてシグナルを伝達します。サブユニットとは、いくつかの分離できるタンパク質から成り立っているタンパク質の一つ一つのタンパク質です。mlgサブユニットはH鎖とL鎖であり、IgαとIgβ二量体はサブユニットはIgαとIgβになります。BCRが凝集(クラスター)すると、ただちにSrcファミリーキナーゼであるLyn、Blk、Fynキナーゼと、チロシンキナーゼであるSyk、Btkを迅速に活性化します。キナーゼとは、リン酸化酵素とも呼ばれ、リン酸基を有するATPなどの分子から別の基質あるいは分子にリン酸を転移する酵素です。続いて細胞内シグナル伝達に必要なシグナロソームと呼ばれる異なった機能をもったタンパク質の複合体の形成が始まります。その複合体(シグナロソーム)にはBCR、チロシンキナーゼや、アダプタータンパク質であるCD19とBLNKなど、さらにシグナル伝達酵素であるPLCγ2、PI3K、Vavなどが含まれます。このシグナロソームから発せられるシグナルは、キナーゼやGTPase、転写因子などの多数のシグナル伝達カスケードを活性化します。その結果、細胞代謝、遺伝子発現、細胞骨格の構成などが変化を受けます。細胞骨格(cytoskeleton,略してCSK)とは、細胞質内に存在し、細胞の形態を維持し、また細胞内外の運動に必要な物理的な力を発生させる細胞内の繊維状構造で、細胞内での各種膜系の変形や移動や細胞小器官の配置に関わります。
BCRシグナル伝達はサイトカインシグナル伝達のメカニズムと同じくらいに極めて複雑すぎるのでありますが、細胞の生存、アポトーシス、増殖、B細胞の抗体産生細胞、メモリーB細胞への分化などの多様な作用をもたらします。核にシグナルが伝わった後の最終的な結果は、細胞の成熟度、抗原の性質、BCRシグナル伝達の強度と持続時間、CD40やIL-21受容体、BAFF-Rなどといった他の受容体からのシグナルによっても大きく影響を受けます。他の多くの膜貫通型タンパク質は、その一部が受容体であり、BCRシグナル伝達の特異的な構成タンパク質に影響を与えます。上の図で示されたCD45、CD19、CD22、PIR-B、およびFcγRIIB1 (CD32) などがBCRシグナル伝達の特異的な構成タンパク質です。
BCRシグナル伝達の強度と持続時間はLyn/CD22/SHP-1経路、Cbp/Csk経路、SHIP、Cbl、Dok-1、Dok-3、FcγRIIB1、PIR-BおよびBCRの内部移行などが影響を与えます。シグナル伝達を抑制する負のフィードバックループによって調節されています。負のフィードバックループ(feedback loop)というのは、フィードバックを繰り返すことで、結果が減少されていくことで、正のフィードバックループは、結果が増幅されていくことです。BCRのような膜型のIgMに抗原が結合して、B細胞が活性化されるには、抗原がBCRを刺激して誘導される細胞骨格の再構築も必要です。サイトカインの受容体シグナル伝達経路を理解するのは最も難しいのですが、それに劣らずB細胞受容体シグナル伝達経路をすべて完全に理解するのは難しすぎますが、さらに勉強した情報をお伝えします。乞うご期待!

上 に抗体の構造と役割の絵図を掲載しておきます。リンパ節でIgM抗体が作られた後に、そのIgM抗体によってオプソニン化されたSARS-CoV-2ウイルスは、補体によってオプソニン化されたウイルスがFDC(Follicular Dendritic Cell)の表面に保持されるようにオプソニン作用を持っている抗体によっても当然保持されます。というのは、FDC(濾胞樹状細胞)は抗体の定常領域(Fc)に結びつくことができる受容体があるからです。オプソニン作用というのは補体と抗体が担う一番重要な作用の1つであることは皆さんご存知ですね。
鼻粘膜や口腔粘膜や上気道の粘膜や下気道の粘膜から侵入したコロナウイルスは、このような粘膜組織に大量に見られる補体や感染初期のみに作られるIgM抗体に捕まえられます。これを補体や抗体によるオプソニン作用といいます。さらに補体や抗体のレセプターを持っている組織に、コロナウイルスを見つけ出す斥候(見張り)として常駐している樹枝状細胞(Dendritic cell、略してDC)に捕捉されます。補体や抗体によってオプソニン化(DCに食いつかれやすくされる味付け作用)された抗原をリンパ節まで運ぶこともご存知ですね。
下のリンパ節の絵図を見てください。T cell Area(Paracortex)とB cell Area(Cortex)の境界あたりに待ち構えているFDCと出会うと、FDCがDCに乗せられてきたオプソニン化されたコロナウイルスの抗原がFDCによって奪い取られるのです。何のためでしょうか?リンパ節の濾胞に存在するB細胞に奪い取った抗原を見せるためなのです。FDCは、補体から奪い取った大量の抗原をB細胞に提示することによってB細胞を活性化させる手助けをするのです。DCの極めて大事な仕事の1つは、オプソナイズされた(味付けられた)Antigenを、リンパ節に運び、FDCに奪い取られる仕事については、免疫学者はしばしば見落としているようです。

しかし、もちろんB細胞が作る抗体が一過性(消失性)のIgMから長期性(生涯性)のIgGを作るために、ヘルパーT細胞(Th)が登場するのです。左の図のリンパ節の絵図の中にT cell Area(Paracortex、訳して傍皮質)と名づけられた青い帯状の部分がありますね。ここがT細胞の存在場所なのです。さらにB cell Area(Cortex、訳して皮質)と書かれた白い部分がB細胞の住処です。B細胞の領域にB細胞がとりわけ多く集まっている部分がリンパ濾胞(Lymphoid Follicle)は体中に張り巡らされているリンパ管の“道の駅”のように体中のあらゆる組織に大小取り交えて500個以上も存在しています。
それではナイーブTh細胞はリンパ節のどの部位でDCによって、どのように活性化され、Tfhに成熟し、かつB細胞をリンパ節のどの部位で出会い、どのようにTfhがB細胞を活性化するのでしょうか? リンパ節で活性化されたTh細胞はTfh(濾胞ヘルパーT細胞)と言います。一言でいうと、まずTh細胞とB細胞とがCognate Antigen(共通認識抗原)と言われる抗原を同時に認識することによってTh細胞のレセプターとB細胞のレセプターとが、まず結合することによってThはTfhになることから始まります。もっと具体的に説明しましょう。
とにかく骨髄で産生されたヘルパーT細胞にしろ、B細胞にしろ、キラーT細胞にしろ、すべての抗原と出会ったことがない上の3つの細胞の前にバージンとかナイーブとかunexperiencedという未経験だという意味の言葉が付きます。一方、抗原と出会ったことのある細胞をexperienced cellといい、抗原に出会う経験をした細胞という意味となります。
B細胞が増えると、濾胞は成長し始め、一次リンパ濾胞は胚中心(germinal center)といわれる二次リンパ濾胞に成長します。B細胞が胚中心(germinal center)で増殖していく時に、共通認識ができる抗原を認識できるヘルパーT細胞から手助けを受けなければ、B細胞はアポトーシスで死んでしまうのです。ここでB細胞に共通抗原を提示してB細胞が自殺するを防ぐのが濾胞ヘルパーT細胞(Follicular helper T cell、略してTfh)なのです。B細胞を自殺から救い出すことができるのです。
重複しますが、骨髄で作られたT細胞やB細胞がどのようにしてリンパ節に運ばれていくのでしょうか? リンパ管から500個以上もあるリンパ節へと寄り道をしながら自分のレセプターにぴったり合う抗原(Cognate Antigen)を探すためにリンパや血液に乗って運ばれています。このようにT細胞やB細胞のレセプターにぴったり合う抗原を英語でCognate Antigenといい、日本語に訳すと共通認識抗原となり、T細胞やB細胞のレセプターの2つが共にぴったり合う共通の抗原を提示してくれるDCを見つけ出す場所が、二次リンパ器官の1つであるリンパ節なのです。これからCognate Antigen(共通認識抗原)という言葉をしばしば使いますからしっかり覚えて下さい。
組織で敵と出会った樹枝状細胞(Dendritic cell、DC)がその敵の抗原をリンパ節に運びます。敵を運ぶ仕事が樹枝状細胞の主要な仕事なのです。これから樹状細胞の名称は正式には樹枝状細胞ですが、今後は樹状細胞で統一します。T細胞やB細胞はCognate Antigen(共通に認識される抗原、つまり共通認識抗原)を求めてリンパ液や血液に乗って体中のリンパ節を漁り回っているのです。これらのリンパ球は一日で体中をひとまわりできると言われます。その途上で立ち寄ったリンパ節でDCが運んでくれるCognate Antigen(共通認識抗原)と出会ってしっかり結びつくと、まず活性化されたB細胞は分裂することによって増殖し、その数をどんどん増やし続けていきます。同じようにB細胞を手助けするヘルパーT細胞(Th)も、ひとたびCognate Antigen(共通に認識される抗原)と出会うと活性化され分裂し、2倍3倍…と増殖し始めます。抗原が二次リンパ器官であるリンパ節にリンパ液や血液に乗って運ばれる形は4つあります。まず1つ目が裸の抗原そのものとして、つまりウイルスそのものとして、2つ目がAPC(抗原提示細胞)に乗せられて、3つ目が補体や抗体によってオプソニン化(味付け化)され、コロナウイルスと戦う様々な組織から運ばれてくるのです抗原として運ばれます。APCであるDCのMHCⅡに抗原のペプチドと結びつけた複合体をThに提示するために運ばれる場合と、あるいはオプソニン化された抗原をFDCによって捕まえさせてB細胞に提示するための2つの場合があることはすでに述べました。復習しておきましょう。DCはヘルパーT細胞に抗原を提示する仕事を、一方FDCはB細胞に抗原を提示する違った仕事をしているのです。混乱しないでください。
上図を見て下さい。リンパ管を流れてきたリンパは、リンパ節に到達すると輸入リンパ管(Incoming Lymphatic)からリンパ節に入っていきます。リンパによって運ばれてきたオプソニン化された抗原はMarginal Sinus(辺縁洞)の穴から通り抜けて、Cortex(皮質)に入り、次にParacortex(傍皮質)入り、補体によってオプソニン化された抗原がリンパ節でB細胞やFDCに認識されなければ、最後はMedullary Sinus(髄洞)から輸出リンパ管を通って素通りしてリンパ節を離れて出ていくのです。Sinusというのは、Cavityとも言い、空洞や穴という意味です。
Marginal Sinus(辺縁洞)の壁は、コロナウイルスがリンパ節に侵入する時に捕まえて貪食しようとする大食細胞で貼り付けられています。大食細胞が貪食してくれるので適応免疫系が処理すべきコロナウイルスの数が減ります。その結果、血流に流れていくコロナウイルスが減る手助けをしてくれます。したがってリンパ節の重要な機能は、リンパに入り込んでいるコロナウイルスを濾過して血流に流さないようにしてくれます。それでは上の図で赤い線の小動脈に流れているT細胞やB細胞はどの部位からリンパ節に入ってくるのでしょうか? それはT cell AreaにあるHEVと書かれている特別な血管壁を通ってT cell Area(Paracortex)からリンパ節にB細胞、T細胞が入りこんでくるのです。HEVについての説明は後で書きます。このT cell AreaはT細胞が蓄積するところであり、輸入リンパ管や小動脈の毛細血管からも流れ出てきたDendritic cell(DC)はこのT cell Areaにもたくさん存在しています。一方、輸入リンパ管から流れ出たB cell Area(Cortex)に集まっています。このB cell Area(Cortex)はリンパ濾胞が存在しているところです。このリンパ濾胞にこそ、リンパ樹状細胞(FDC)がB細胞にDCが持ってきたオプソニン化した抗原を見せるためにリンパ濾胞を住所としているのがFDCです。言い換えると、リンパ節はAPCやTリンパ球やBリンパ球やマクロファージや、かつオプソニン化された抗原を満載しているFDCの5つの免疫のプレイヤーが集まる場所なのです。しかも、この5者のプレイヤーともすべて同じCognate Antigen(共通認識抗原)を認識することができるお互いに協力し合う仲間と言えます。
リンパ濾胞でB細胞がFDCによって提示されたCognateAntigen(共通認識抗原)を見つけ出すとBCRのレセプターとFDCによってオプソニン化されたCognateAntigen(共通認識抗原)の複合体は一緒にB細胞の細胞内に取り込まれます。ひとたびB細胞に取り込まれてしまうやいなやCognateAntigen(共通認識抗原)はB細胞によって酵素的に消化されてMHCⅡ分子の上に乗せられてT細胞に見せるためにB細胞の細胞膜に提示されます。しかしながらB細胞が十分に成熟するためにはB細胞はFDCからCognateAntigen(共通認識抗原)をもぎ取った後、T細胞の持つCD40Lのタンパクの手助けが必要なのです。このCD40LこそB細胞を刺激するCo-stimulation(補助刺激)となることはすでに述べました。このTh細胞のCD40LのCo-stimulation(補助刺激)はB細胞の表面に存在するCD40と結びつくのです。Th細胞だけがこのCo-stimulation(補助刺激)をB細胞に提供することができるのですが、一方Th細胞の方もAPCであるB細胞が自分に提示するCognateAntigen(共通認識抗原)がなければ十分に成熟できないのです。皆さんはB細胞は抗体を作るだけの仕事が多いと思いですが、実はB細胞はMHCⅡ分子を持っているAPCの仕事もこのリンパ濾胞で発揮できるのです。Bリンパ球はFDCからCognateAntigen(共通認識抗原)を奪い取る一方で、今度は奪い取ったこのCognateAntigen(共通認識抗原)をThに見せることによってThをTfhまで成熟させることができるのです。ややこしいでしょう。しかし極めて面白いでしょう。こういう離れ業ができる場所を提供しているのが、リンパ節のリンパ濾胞であり、最終的にはこのリンパ濾胞の中でコロナウイルスに対して効果的な抗体を作れるBリンパ球を増やすための成熟しきった特定の場所がリンパ濾胞の胚中心(germinalcenter)なのです。仕事が終われば胚中心も消えてなくなるのです。
さらにTh細胞がB細胞を助けるのみならず、Th細胞の方もB細胞の手助けが必要なのです。それはTh細胞がTfh細胞まで成熟するにはB細胞がもつB7タンパクとB細胞膜の上にあるICOSLタンパクの手助けが必要なのです。このB細胞のB7タンパクと未熟なTh細胞のCD28と結びつくと同時にB細胞のICOSLタンパクは同時にTh細胞のICOSタンパクと結びつく必要があるのです。ICOSは英語で Inducible Costimulatorであり、Inducible(誘導される)のIと Costimulator のCosをとってICOSと名づけたのです。日本語で訳すと、誘導補助刺激分子となります。Costimulatorは共刺激分子や副刺激分子と訳されることが多いのですが、正しくは補助刺激分子です。
非常に詳しく書きすぎたので整理しましょう。まず、Cognate Antigen(共通認識抗原)を持ったナイーブT細胞とナイーブB細胞が最初にリンパ節のT cell Area(Paracortex)とB cell Area(Cortex)で出会いますと1時間かかって、形質細胞になったいくつかのB細胞は増殖し、初期の親和性の低いIgM分泌抗体を生み出します。もちろん、1時間の出会いでこのような形質B細胞はクラススイッチやSomatic hyper-mutationされることはありませんが、初期の感染に対して素早く対抗できます。他の形質細胞にならなかったB細胞と境界でThはリンパ濾胞へと一緒に移動します。そこでB細胞とThはお互いに助け合いながらTh細胞とB細胞は、Th細胞の上にあるCD40LとB細胞のCD40が結びついて相互に手助けしながら、かつB細胞のMHCⅡとB7とICOSLはTh細胞を手助けしてTh細胞をTfhに成熟し、さらにどんどん増えていくB細胞は一次リンパ濾胞を二次リンパ濾胞に変えて、つぎに二次リンパ濾胞の一部を胚中心(germinal center)に変えて、最後は増えすぎたB細胞が胚中心(germinal center)の一部をダークゾーン(暗帯とか、暗領域)に変えてしまいます。この胚中心(germinal center)のダークゾーン(暗領域)でクラススイッチと体細胞超変異(Somatic hyper-mutation)が起こるのです。
ここでヘルパーT細胞(Th)とB細胞(B cell)との関係をまとめておきましょう。最終的にThとB cellはどんな分子によって結合することによってクラススイッチをし、体細胞突然変異を起こすのかをまとめましょう。全部でThとB cellのそれぞれ5種類の分子同士が結びつく必要があります。1つ目がThのTCRとB cellのMHCⅡに乗せられたペプチド、2つ目がThのCD40LとB cellのCD40、3つ目がThのCD28とB cellのB7、4つ目がThのICOSとB cellのICOSLの4つです。
さてヘルパーT細胞の手助けがなければメモリーB細胞ができないのでしょうか?
B cellがメモリーB cellになるためにはThのCD40LがB cellのCD40に結びついてThからCD40Lという co-stimulation molecule(補助刺激分子)の手助けが絶対に必要であるからです。なぜなのかはまだ解明されていませんが、実験的に知られている事実です。逆にメモリーT cellが生まれるためにはB cellのB7がT cellのCD28に結びつかなければ生まれないのではないかと考えています。というのはB7はT cellに対してはco-stimulation moleculeとなっているからです。メモリーT cellは、T cellのCD28にB cellのB7が結びついたという経験を記憶しているT cellが永続して生き続けていると考えています。しかもB cellのみならず、全てのAPCが発現していますから、機会あるごとにこのメモリーT cellは非特異的に刺激され続けるので、メモリーT cellとして生き続けるのではないかと考えています。と同時に、このメモリーT cellは特異的な抗原を認識したT cell receptorに結びついた刺激を、決まり切ったFc部分のIgMというFcの遺伝子をIgGのFcの遺伝子に置き換えるだけであります。ところがハイパーミューテーションはクラススイッチをした後のFabの遺伝子のアトランダムな組み替えによって敵と結びつく親和性がある閾値を超えてしまうほどの強い親和性を持つほどのFabを持つようになったT細胞がメモリーT cellとして永遠に生きるのではないかと考えています。
共刺激とは何でしょうか?なぜ共刺激は非常に大切なのでしょうか?
共刺激は英語でco-stimulation といい、その共刺激を与えることができる分子をco-stimulation moleculeといいます。co-stimulation moleculeの正しい訳は副刺激です。抗原提示細胞(APC)に抗原提示されるだけの一次的な刺激だけではTリンパ球は活性化できないのです。さらに免疫応答を実行するためには適応免疫細胞が必要とする非特異的な二次シグナル(co-stimulation signal)が絶対に必要なのです。APCの表面にある非特異的なB7というタンパクがT細胞のCD28に結びつく必要があるのです。さらにT細胞が増殖し分化し成熟し生存をし続けるためにはもう1つ別の補助刺激がBリンパ球のもつICOSLであります。このICOSLに対するT細胞の補助刺激受容体がICOSであります。ICOSは英語で Inducible Costimulatorであり、Inducible(誘導される)のIと Costimulator のCosをとってICOSと名づけたことは既に述べました。このBリンパ球のICOSLとTリンパ球のICOSの関係はどちらもリンパ球であるので適応免疫同士の関係と思われますが実は間違っています。なぜならば、Bリンパ球のICOSLはBリンパ球がAPCとしての仕事としてICOSLを用いているのです。したがってICOSLを stimulation 分子として言うべきであるのですが、免疫学者はこの意味を理解していないようです。残念ですが。アハハ!
T細胞に必要な補助刺激がなくなればどうなるでしょうか?
T細胞の増殖、分化、生存に必要なのはAPC(樹状細胞や大食細胞)が提示する抗原を認識するだけではなく、APCの持つB7というタンパク質の補助刺激であります。この補助刺激(co-stimulation)によってT細胞が刺激されなければ、このT細胞はアネルギーという状態となり、死んでしまいます。生き続けて仕事をするためには、補助刺激のB7が絶対に必要です。
または免疫寛容の発達につながる可能性があります。 B細胞共刺激 後者の場合、抗原特異的Th2細胞またはTfh細胞による認識が誘導され、TCRがMHC-抗原複合体に結合することでB細胞が活性化されます。その後、Th2細胞上でCD40L(CD154)が合成および提示され、B細胞上のCD40に結合するため、Th2細胞はB細胞を共刺激することができます。この共刺激がなければ、B細胞はそれ以上増殖できません。B細胞の共刺激は、補体受容体によって代替的に提供されます。微生物は補体系を直接活性化し、補体成分C3bが微生物に結合する可能性があります。C3bがフラグメントiC3b(C3bの不活性誘導体)に分解された後、C3dgに切断され、最後にC3dに切断され、微生物表面に結合し続けます。B細胞は補体受容体CR2(CD21)を発現して、iC3b、C3dg、またはC3d。この追加の結合により、B細胞は抗原に対して100〜10,000倍感受性が高くなります。成熟B細胞上のCR2は、CD19およびCD81と複合体を形成します。この複合体はB細胞補助受容体と呼ばれます抗原に対するそのような感受性増強のための複合体。アプリケーション アバタセプト(オレンシア)は、関節リウマチの治療に承認されたT細胞共刺激モジュレーターです。サイトカイン活性化T細胞によって分泌されるが、両方と考えられる開始および関節リウマチに関連した免疫学的に駆動される炎症を伝播します。可溶性融合タンパク質であるオレンシアは、完全なT細胞の活性化に必要な共刺激シグナルを変化させることによって機能します。Belataceptは、腎移植で使用するための拒絶反応抑制薬としてテストされている別の新しい分子です。 新しい共刺激超アゴニスト 薬であるTGN1412は、ロンドンのノースウィックパーク病院での臨床試験の対象でした。6人のボランティアが薬を与えられてから数分以内に重病になったので、裁判は論争に巻き込まれました。 本質的に、共刺激分子は、T細胞と相互作用する「点滅する赤色光」として機能し、樹状細胞材料によって提示されている材料が危険を示していることを伝えます。抗原を提示しながら共刺激分子を表示する樹状細胞は、T細胞を活性化することができます。対照的に、共刺激分子を表示しない樹状細胞によって提示された抗原を認識するT細胞は、一般にアポトーシスに駆り立てられるか、または抗原との将来の遭遇に反応しなくなる可能性があります。

そ れでは上絵図に B cellとTh cellがどのような分子同士と結合して相互に刺激しあっていたか、またしているかを一目でわかるように示しました。B cellが提供する分子は上からMHCⅡとB7とICOSLとCD40の4つです。Th cellが提供する分子は上からTCRとCD28とICOSとCD40Lの4つの分子です。ICOSは英語でinducible co-stimulatorとかinducible T-cell co-stimulation molecule で略してICOSといい、日本語では誘導性補助刺激分子と訳します。ICOSLは英語でinducible co-stimulator ligandで略してICOSLです。 co-stimulatorという英語の正しい訳は補助刺激分子です。
B cellとTh cellはそれぞれ4種類の分子同士の結合は同時でなくてもいいのですが必要です。刺激さえすればいいのです。その内ThがB cellを補助する(助ける)結びつきは2つあります。1つ目がThのTCRがB cellのMHCⅡと黄色い丸印のペプチド複合体と、2つ目がThのCD40LがB cellのCD40と結合して刺激してTh cellがB cellをヘルプ(補助)しています。これらの結合はずっと結びついているのではなくて刺激されれば離れてもいいのです。いや、離れるべきなのです。そうでなければそれぞれの細胞が自由に動き回ることができなくなるからです。
次にB cellがTh cellを刺激する手助け(補助)をする結びつきは、1つ目がB cellのMHCⅡと黄色い丸印のペプチド複合体(抗原)をTh cellのTCRに提示することによって刺激して、T細胞はAPCであるB細胞に敵を認識させられています。皆さん、B cellは抗体を作る専門家であるはずなのにDC(樹状細胞)と同じようにMHCⅡタンパクにペプチド複合体を作り、Th cellに抗原である黄色い丸印のペプチドをMHCⅡに乗せて提示することができるのは不思議に思いませんか?実はB細胞もAPCの一つなのです。それはMHCⅡを持っている細胞がAPCの定義であり、B cellもMHCⅡ分子を持っているのでAPC(antigen presenting cell、訳して抗原提示細胞)の1つであるからです。実はB cellは4つの仕事ができます。1つ目が抗体産生工場であり、2つ目が抗原提示細胞(APC)でもあり、3つ目がB cellのB7がTh cellのCD28に結びつくことでB cellはTh cellを刺激する手助け(補助)をしているのです。4つ目は、B cellのICOSLがThのICOSに結びつくことによってB cellはThに対する補助刺激分子となっているのです。このようにリンパ節の胚中心において役割は異なりますが、Th cellとB cellがそれぞれ相互に刺激し合い、補助し合って、IgG抗体産生のみならず、メモリーB cellとメモリーヘルパーT cellとメモリーキラーT cellを作り出しているのです。その相互刺激作用のための結びつきはTCRとMHCⅡと黄色い丸印のペプチドの複合体が出発点となっているのです。つまりT細胞の受容体であるTCRに抗原提示細胞(APC)のMHCⅡ分子に乗せられた病原体のペプチド断片を乗せて、この敵をT細胞に見せることによりT細胞が活性化されることからすべての相互作用が始まるのです。ややこしいですが、いかに人体から免疫が守っているかがお分かりになるでしょうか?にもかかわらず、残念なことにこの世にありもしない自己免疫疾患を作り立てて病気づくりに専念している医薬業界に激しい憤りを感じるのは間違いでしょうか?アハハ!
以上を述べたようにThとB cellは助けてもらったり、助けたりして、結局は相互に助け合うことによって、最終的にはB cellが抗体のクラススイッチをし、かつ体細胞突然変異を行うことが可能となるのみならず、メモリーB cell、メモリーヘルパーT cell、メモリーキラーT cellの全ての免疫記憶細胞を生み出しているのです。皆さん、全部理解し、覚えられますか???
それではどのようにしてキラーT細胞が活性化され、メモリーキラーT細胞が生まれるのでしょうか?基本的には今まで説明してきたメモリーB cellが形成されるのと同じ原理ですから、理解は容易いと思います。アハハ!すいません。
既に述べたのですが、メモリーキラーT細胞が生まれるためには、3つの大食細胞や樹状細胞やB細胞などの抗原提示細胞(APC)に直接にウイルスが感染しなければ、絶対に記憶キラーT細胞は生まれないのです。なぜならば、APCに感染しない限りは、ウイルスが極めて危険な病原体であるという証拠となる人間が持っていなくてウイルスなどが持っている独自のRNAやDNAや、さらにこれらの遺伝子が作らせる人間が持っていないタンパクを危険な異物として38億年かけて人類の免疫の遺伝子に覚え込ませた敵の特徴があります。この特徴をPAMP(Pathogen-associated molecular patterns、訳して病原体関連分子パターン)敵として認識するPRR(pattern-recognition receptor訳して、パターン認識受容体)が認識されないからです。さらに病原体と戦った後、傷害を受けて細胞に見られる独特なもう1つDAMP(damage-associated molecular pattern 、訳して傷害関連分子パターン)という病原体との戦いの証拠がPAMPと同じく遺伝子で記憶されているDAMPについても後で詳しく説明します。
それではまずPAMPとは何でしょうか?なぜそんなに大事なのでしょうか?それは自己免疫性疾患がないという証拠の1つとなるからです。38億年の免疫の進化において、自己の遺伝子が成分を異物と認識したことはないにもかかわらず、現代の医学は敵として認識する必要がないどころか人間の生命を存続させてくれる自己の成分を攻撃する自己免疫は存在し得ないにもかかわらず、自己免疫性疾患を捏造して資本主義医療を恥ずかしげもなく展開し続け、病気を作り続けています。アハハ!この笑いは皮肉で悲しい笑いです。アハハ!
自然免疫における病原体の認識 PRRのレパートリーは非常に広範であり、同様に、PRRによって認識される病原体のクラスは非常に多様です。なぜ自然免疫だけがPRRを持っているのでしょうか?それは後天免役はPRRという自然免疫の働きによって敵を認識してもらって、初めてその敵を殺すなり共存するなりの処理を効率よくするだけの仕事をしているだけです。病原体などの敵を認識してくれるのは自然免疫しかないからです。
産まれ持った自然免疫しかできない病原体認識の中心的な特徴は、生化学的組成がまったく異なり、ライフサイクルもまったく異なるウイルスや細菌などの微生物が、宿主である人間のPRRによる様々の類似したメカニズムによって認識されることです。さらに、このPRRのシステムの重要な特性は、単一の種類の病原体が1つのタイプのPRRによってのみ感知されることはないということです。むしろ、多くの異なるPRRがさまざまなPAMPを介して特定の病原体を認識することができるので、その認識に基づいて反応する適応免疫が迅速で強力な戦いを始めることができ、その炎症反応の特異性も可能になります。
この世の病原体で一番強敵なのは、細胞に感染せざるを得ないウイルスなのです。SARS-CoV-2はまさに世界中を大混乱に陥れているのはウイルスであるからです。ウイルスを認識したPRR活性化は、感染を効率的に除去する抗ウイルス免疫で応答するのですが、残念ながら人間の免疫よりもウイルスの方が遥かに狡猾であるので殺すことができないのみならずワクチンもできないのです。
ウイルスは、表面糖タンパク質、DNA、RNAなど、構造的に多様なPAMPを多数持っています。これらの免疫を刺激してくれるDNAやRNAなどのヌクレオチドは、感染性ビリオンのみならず、ウイルスの遺伝子複製中やビリオン複製中に産生されます。宿主は広範囲のウイルスのヌクレオチドを認識できるセンサーを保有しているので、ウイルスDNAはTLR9とDAI(DNA-dependent activator of IFN-regulatory factorsで、訳してDNA依存性インターフェロン調節因子活性化分子)によって認識されますが、ssRNAはTLR7とTLR8によって検出され、最後にdsRNAと5′-三リン酸RNAがRLR、TLR3、PKRを活性化します。ssRNAは一本鎖のRNAで、dsRNAは二本鎖のRNAです。さらに、いくつかのウイルス糖タンパク質がTLR2およびTLR4によって認識されます。例えば、RSV(Respiratory syncytial virus、訳して呼吸器合胞体ウイルス)の融合タンパク質はTLR4を活性化します。TLR2は単純ヘルペスやサイトカインウイルスなどを含む様々なウイルスによって活性化されます。さらにTLR2は麻しんウイルスのヘマアグルティニン(hemagglutinin)などの成分によって活性化されます。免役応答の開始はウイルスのPAMPがその受容体であるPRRに結合するところから開始されるのです。
単純ヘルペス(HSV)のPAMPがどのように自然免疫のPRRによって認識されるのでしょうか?
HSVは、HSV-1とHSV-2の2つのタイプがあり、どちらもエンベロープをもったDNAウイルスです。この単純ヘルペスのHSV-1とHSV-2のウイルスは、歯肉口内炎、口唇ヘルペス、性器ヘルペスから、重篤で致命的な脳炎、髄膜炎、新生児ヘルペスまで様々な感染症を引き起こします。HSV感染中、細胞の種類とウイルス複製サイクルの段階に応じて、PRRによる病原体認識の複数のメカニズムが機能しています。まず、HSVビリオンまたはビリオン表面糖タンパク質が細胞表面のTLR2と相互作用します。HSVによるTLR2の活性化は、サイトカイン産生をもたらし、入ってくるウイルス粒子によって誘発される他の初期応答は、ウイルスDNAによって誘導され、TLR9によって媒介される強力なI型IFN(IFN-α、 IFN-β)を産生しますが、この産生はpDC(plasmacytoid dendritic cells、訳して形質細胞様樹状細胞、略してpDC)だけに限定されます。pDC以外の細胞型もIFN-α、IFN-βと結合するとⅠ型IFNを産生します。HSVは複数の細胞認識システムによって検出されます。これらのシステムは、細胞の種類や時間に依存して動作し、抗ウイルス反応を引き起こし、IFN-αやIFN-βなどのⅠ型IFNを産生します。
病原体関連分子パターン(PAMP)は、微生物の生存に絶対不可欠なタンパクとして保存されている小分子モチーフです。それらは、植物と動物の両方で、トール様受容体(TLR)と他の様々なパターン認識受容体(PRR)によって認識されます。グリカン(多糖体)や複合糖質など、さまざまな種類の分子がPAMPとして機能します。PAMPの代表であるグラム陰性細菌が持っているリポ多糖(Lipo-poly-saccharide、略してLPS)は、自然免疫系の認識受容体であるTLR4によって特異的に認識されます。他のPAMPは、細菌の持つフラジェリンはTLR5によって認識され、グラム陽性細菌の細胞壁に見られるリポテイコ酸(Lipo-teichoic acid、略してLTA)はTLR2によって認識され、細菌の細胞壁にあるペプチドと糖からなる高分子化合物ペプチドグリカン(Peptidoglycan)はTLR2によって認識されます。さらにTLR3によって認識される二本鎖RNA(dsRNA)やTLR9によって認識される非メチル化CpGモチーフなどがPAMPとしてPRRに認識されるのです。
PRRによるPAMPの認識の結果、インターフェロン(IFN)が産生され、宿主免疫細胞におけるいくつかのシグナル伝達カスケードの活性化を引き起こします。結核などを引き起こすマイコバクテリアは、宿主マクロファージで生き残る細胞内細菌として有名です。マイコバクテリアの壁は脂質と多糖類で構成されており、ミコール酸も大量に含まれています。なぜ結核菌は大食細胞が殺せないのか? ミコール酸(mycolicacids)は、結核菌が菌体の最外周部に持っている分子量の大きな脂肪酸の総称で、ミコール酸はいつかの脂肪酸の集まりで純物質ではないのです。多種類の脂肪酸の集まりであるミコール酸の炭素数はだいたい60個から90個程度であり、シクロプロパンを構造に含んでいます。結核菌は増殖する時にミコール酸を合成し、結核菌の菌体の最外周部をミコール酸で囲み、結核菌のペプチドグリカンが主要成分の細胞壁よりもさらに外側にあります。アラビノガラクタンが主成分の部分よりもさらに外側にミコール酸はあって、自身を保護しています。ペプチドグリカン(Peptidoglycan)は、細菌の細胞壁によくあるペプチドと糖からなる高分子化合物の一種であり、狭義にはムレイン酸(murein)としても知られ、真正細菌の細胞膜の外側に層を形成する細胞壁の主要物質です。アラビノ-ガラクタン-プロテイン(AGP)は植物組織に普遍的に分布しているプロテオグリカン(タンパク多糖体)で,主に細胞壁(細胞外マトリックス)に局在しています。AGPは一般的にはヒドロキシプロリン(Hyp)に富むコアタンパク質に,ガラクトース(Gal)とL-アラビノース(L-Ara)に富むアラビノガラクタン(AG)糖鎖が結合しているのがアラビノ-ガラクタン-プロテイン(AGP)です。この構造がマクロファージに貪食され取り込まれても内部で生き続けられる原因の1つとなっているのです。なお1分子のミコール酸で1つの結核菌を囲んでいるわけではなく、ミコール酸同士はトレハロースを間に挟んで連なっている上に、ミコール酸は内側にあるアラビノ-ガラクタンとも結合しているのです。マイコバクテリアの精製されたこのような細胞壁成分は、主にTLR2とTLR4を活性化します。リポマンナンとリポアラビノマンナンは、強力な免疫活性化リポグリカンです。TLR1と結合したTLR2は、結核菌由来の細胞壁リポタンパク質抗原を認識できます。これは、マクロファージによるサイトカインの産生も誘導します。さらにTLR9はマイコバクテリアDNAによっても活性化されます。このように非自己成分であるタンパクや他の有機成分などは自然免疫のAPCのTolllikereceptor(TLR)によって認識されて危険が差し迫っているというシグナルを獲得免疫(適応免疫)に伝えるのです。自己免疫性疾患はこのようなシステムを完全に無視した架空の病気であることを死ぬまで証明し続けるために医療を続けたいのです。自己免疫性疾患は医薬業界が作り上げたバーチャルフェイクです。どうせ強大な医薬業界に潰される覚悟はしていますがいつになるでしょうかね?アハハ!
ダメージ関連分子パターン(damage-associated molecular patterns、略してDAMP)は、外傷または病原体による感染によって損傷または死にかけている細胞から放出される自然免疫応答の構成要素である細胞内の分子です。これらの分子は、人体が病原体によって細胞に損傷が起こったり、あるいは感染が起こっていることを免疫に知らせる警告サインとして機能するため、危険関連分子パターン、危険信号、およびアラーミン(alermin)としてもDAMPは知られています。DAMPは、外傷や病原体による細胞への損傷に応答して細胞外に放出される細胞の内側に生じている危険を細胞外の免疫に知らせる危険信号です。DAMPが細胞から放出されると、DAMPがパターン認識受容体(PRR)に結合することにより、非感染性の敵との戦いが終わった処理をするための炎症反応を促進します。炎症は、患部から有害な侵入者を取り除くのみならず、治癒プロセスを開始することにより、生物への将来の損傷を軽減するために使用されるため、自然免疫応答の重要な側面です。にもかかわらず、現代の医療を炎症を目の敵にしているのは非常に残念です。
例えば、サイトカインIL-1αは細胞の核内で発生するDAMPであり、細胞外に放出されると、PRRのIL-1Rに結合し、PRR IL-1Rが炎症反応を開始します。外傷の後始末のために放出されたIL-1αというDAMPは非感染性炎症反応であり、一方、病原体関連分子パターン(PAMP)は、感染性病原体誘発性炎症反応を開始させ敵を殺すまで永続化します。多くのDAMPは、組織の傷害が起こった後、細胞の外に放出される明確な機能を持っていた核タンパクや細胞質にあるタンパクなのです。細胞内から細胞外の排泄は、DAMPを細胞内の還元環境から細胞外の酸化環境に移動させ、それらの機能的な変性を引き起こされ、DAMPの成分の機能も失われます。核および細胞質ゾルのDAMP以外にも、ミトコンドリア、細胞内顆粒、細胞外マトリックス、小胞体、原形質膜など、さまざまな部位に由来する他のDAMPがあります。DAMPの主要な受容体は、やはりPAMPと同じでTLRです。
DAMPの出所部位と主なDAMPとそのDAMPの受容体の一覧表
| 出所部位 |
| 主要なDAMP | 受容体 |
| 細胞外マトリックス |
| ビグリカン | TLR2、TLR4、NLRP3 |
|
|
| デコリン | TLR2、TLR4 |
|
|
| バーシカン | TLR2、TLR6、CD14 |
|
|
| LMWヒアルロン酸 | TLR2、TLR4、NLRP3 |
|
|
| ヘパラン硫酸 | TLR4 |
|
|
| フィブロネクチン(EDAドメイン) | TLR4 |
|
|
| フィブリノーゲン | TLR4 |
|
|
| テネイシンC | TLR4 |
| 細胞内コンパートメント | サイトゾル | 尿酸 | NLPR3、P2X7 |
|
|
| S100タンパク質 | TLR2、TLR4、RAGE |
|
|
| HSP(熱ショックタンパク質) | TLR2、TLR4、CD91 |
|
|
| ATP | P2X7、P2Y2 |
|
|
| F-アクチン | DNGR-1 |
|
|
| シクロフィリンA | CD147 |
|
|
| Aβ | TLR2、NLRP1、NLRP3、CD36、RAGE |
|
| 核 | ヒストン | TLR2、TLR4 |
|
|
| HMGB1 | TLR2、TLR4、RAGE |
|
|
| HMGN1 | TLR4 |
|
|
| IL-1α | IL-1R |
|
|
| IL-33 | ST2 |
|
|
| SAP130 | ミンクル(Mincle) |
|
|
| DNA | TLR9、AIM2 |
|
|
| RNA | TLR3、TLR7、TLR8、RIG-I、MDAS |
|
| ミトコンドリア | mtDNA | TLR9 |
|
|
| TFAM | レイジ |
|
|
| ホルミルペプチド | FPR1 |
|
|
| mROS | NLRP3 |
|
| 小胞体 | カルレティキュリン | CD91 |
|
| 顆粒 | ディフェンシン | TLR4 |
|
|
| カテリシジン(LL37) | P2X7、FPR2 |
|
|
| EDN(好酸球由来ニューロトキシン) | TLR2 |
|
|
| グラニュライシン | TLR4 |
|
| 原形質膜 | シンデカン | TLR4 |
|
|
| グリピカン | TLR4 |
皆さん、私がなぜこんなにDAMPにこだわっているのかお分かりですか?既に私はアルツハイマーの原因はアミロイドβと世界中の医薬業界は思い込み続けており、アミロイドβを減らす薬を何十年も何十兆円もかけて失敗に終わっていますが、なぜかというのはすでに書きました。ここを読んで下さい。アミロイドβはあくまでも海馬の細胞に感染したヘルペスと免疫が戦った後の傷害の産物に過ぎないのです。つまり、DAMP(Damage-associated molecular patterns)と名づけられるダメージ(損傷)関連分子パターンの1つがアミロイドβなのです。すでに述べたように、ヘルペスによる細胞死や細胞の損傷など,ヘルペスによって細胞が崩壊した後、放出されたものであり,細胞の危機を免疫に知らせるアラームとして機能しているだけであり、決して自己の免疫が自分の細胞を攻撃した結果ではないのです。これまでに報告されているDAMPsの一覧表をタンパク質から核酸まで多岐にわたる様々な分子を掲載したのです。
今朝の3月10日の朝日新聞に大スクープよろしく「潰瘍性大腸炎」 患者9割に特定の「抗体」 京大発見 の特ダネが出ていましたが、潰瘍性大腸炎はすでに述べたように決して自己免疫性疾患ではなくて、化学物質をIgGで戦うか、ヘルペスと戦うかによって生じる腸疾患に過ぎないのです。ここを読んで下さい。その新聞の内容を掲載しておきましょう。
京都大の研究グループが、原因不明の下痢や血便を繰り返す難病「潰瘍(かいよう)性大腸炎」の患者の9割に、特定の「抗体」があることを見つけたと発表した。抗体を測る検査キットを企業と開発し、新たな診断法にしたいとしている。 潰瘍性大腸炎には自己免疫がかかわると考えられている。自己免疫とは、免疫反応でできる抗体が、誤って自分の体内にもともとある物質を攻撃する現象だ。抗体は本来、病原体を攻撃する。 そこでグループは患者112人の血液で、自分の体内にある物質に反応する「自己抗体」を調べた。その結果、患者の9割に「インテグリンαVβ6」というたんぱく質に対する抗体があった。他の病気の患者には、この抗体は少ないこともわかった。 この抗体は、症状が悪化すると増え、改善すると減る傾向があり、病気の診断や病状の判断に役立つ可能性がある。グループの桑田威医員は「病気の原因解明や治療法開発につながる可能性もある」と話している。
それではインテグリンαvβ6とは何でしょうか?
ケラチン細胞のリガンドであるフィブロネクチンおよびビトロネクチン上の移動に必須であるタンパク質です。αvβ6. インテグリン(β6)は、正常組織ではほとんど発現がみられないが、上皮修復や腫瘍形成の過程で発現が増加し、主に間質を構成するフィブロネクチンと結合することが知られている。インテグリンαVβ6というタンパク質に対する自己抗体が潰瘍性大腸炎患者の約90%に認められることを発見したと京都大学は発表しましたが、これは自己抗体ではなくて大腸の細胞が崩壊した後の細胞傷害関連分子であるDAMPであるインテグリンαVβ6をB細胞が処理するときにインテグリンαVβ6をオプソニン化する時に作ったIgM抗体にすぎないのです。この抗体はMALTによって作られた二量体のIgA抗体であります。この二量体のIgA抗体は排除の抗体であり、殺しのIgMやIgG抗体ではないのでこの抗体に対する抗体を作っても意味がないのです。
ダメージ関連分子パターン(DAMP)は、自然免疫系を刺激して敵を認識させて適応免疫に危険を伝えるのです。DAMPは、細胞の種類や上皮組織または間葉組織などの損傷した組織によってヘルペスとの戦いでダメージを受けた分子の違いによって様々のパターンがありますが、すべて生物内の自然免疫応答を刺激するという共通の特徴を共有しています。タンパク質DAMPには、熱ショックタンパク質やHMGB1 などの細胞内タンパク質、およびヒアルロン酸フラグメントなどの組織損傷後に生成される細胞外マトリックスに由来する材料が含まれます。 HMGB1とは、高移動グループボックスタンパク質1、英語でHigh mobility group box protein 1、略してHMGB1ですが、インフラマソームの活性化時にグリア細胞とニューロンによって放出されるどこにでも存在する核タンパク質であり、標的細胞上の終末糖化産物受容体(receptor for advanced glycation end products、略してRAGE)およびトール様受容体(TLR)4の受容体を活性化します。RAGEとは、私たちの細胞は細胞表面にある受容体でこのAGE修飾を認識します。AGEとは、終末糖化産物(advanced glycation end productsで、略してAGE)です。
それではDAMPを非タンパク質とタンパク質に分けながら個々に詳しく説明し続けましょう。
非タンパク質DAMPには、1)ATP、2)尿酸、3)ヘパラン硫酸および4)DNA、5)RNA、6)単糖類、7)多糖類、8)プリン代謝物、9)アデノシンが含まれます。一方、タンパク質DAMPには、1)HMGB1、2)DNAとRNA、3)S100タンパク質、4)単糖類と多糖類が含まれます。
まず非タンパク質から説明しましょう。
DNAとRNAが腸管の微生物であるウイルスや細菌と免疫細胞が戦った時に、殺された微生物から漏れ出たDNAやRNAが核以外またはミトコンドリア以外の場所に見つけられ、DAMPとして大食細胞のTLR9や細胞質に存在するDAI(英語でDNA-dependent activator of interferon-regulatory factors、訳してDNA依存性IRF活性化因子、略してDAI)によって認識されると、残党のヘルペスや細菌などを処理するために、さらに他の免疫の細胞の活性化と免疫反応を促進させ、敵を皆殺しにします。
ヒアルロン酸は線維芽細胞や滑膜細胞などで産生されています。この線維芽細胞や滑膜細胞に感染したヘルペスがIFN-αやIFN-βやNK細胞やキラーT細胞などで細胞もろとも殺された時に、DAMPとして細胞外マトリックスにヒアルロン酸分解産物として大量に放出されると、マクロファージや樹状細胞のトール様受容体2(TLR2)、TLR4、またはTLR2とTLR4の両方を介して炎症シグナルを伝達して、さらにヘルペスを殺そうとするのです。したがってDAMPというのは、細胞がダメージを受けて崩壊した証拠であると同時に、そのDAMPを作り出す敵をさらに処理するためにますます免疫反応が盛んになるのです。特に最後に残された細胞に感染する病原体はヘルペスですから、そのヘルペスをできる限り隣接する細胞にさらに感染し続けることを抑えるために存在しているのがDAMPなのです。すでに書きましたが、アルツハイマーの患者に見られるアミロイドβはまさに脳の海馬の細胞に感染したヘルペスを処理するために細胞をヘルペスもろとも殺しきった結果、崩壊した細胞から漏れ出た代表的なDAMPの産物であるにも関わらず、世界中の優れた医者たちはアミロイドβはアルツハイマーの原因と言い続けているのです。抗ヘルペス剤を大量にアルツハイマーの患者に投与し続けばアルツハイマーの症状が初期に制御できるのに残念ですね。ヒアルロン酸(Hyaluronicacid)は、細胞外マトリックスの主成分であるのですがDAMPとしてのヒアルロン酸やヒアルロン酸分解産物はマクロファージや樹状細胞のトール様受容体2(TLR2)、TLR4、またはTLR2とTLR4の両方を介して炎症シグナルを伝達して、ヒアルロン酸が最も多い皮膚のマトリックスの創傷修復のための組織再生、炎症反応、血管新生に重要な役割を果たしているのです。TLRとDAMPとしてのヒアルロン酸やヒアルロン酸分解産物は自然免疫において様々な組織の細胞にヘルペスが感染しているという警告を発する重要な役割を果たしているのです。DAMPが見つかれば抗ヘルペス剤を投与すれば全て原因治療となり、この世から不明な病気や自己免疫性疾患は消えてしまうのですが、世界中の医者は誰も気づいていないのです。
非タンパク質DAMPであるプリン代謝物やヌクレオチドであるATPやヌクレオシドであるアデノシンが細胞崩壊と共に壊れた核から細胞外(マトリックス)空間に到達すると、プリン作動性受容体を介してシグナル伝達することにより、細胞にヘルペスが感染しているというメッセージを免疫系に伝える危険信号として機能しているのです。ATPとアデノシンは、ヘルペス性壊死性の細胞死が生じる時に、細胞の壊滅的な破壊の後に高濃度で放出されます。アデノシンはP1受容体(purinergicreceptorで、略してP1)で、アデノシンをリガンドとする一群の細胞表面受容体で、Gタンパク質共役型受容体(GPCR)に属し、A1、A2、A2BおよびA3に分類されます。ATPとアデノシンはあくまでもDAMPによる細胞の崩壊によって漏れ出た屑(くず)であり、決して自己免疫性疾患によって攻撃される自己の成分ではないのです。あくまでもヘルペスウイルスが感染した細胞がヘルペスと共に壊死してしまった後の細胞の部品であって、その部品を免疫が処理するために攻撃したとしてもそれは大食細胞によって貪食されてしまい、跡形も消え去ってしまうゴミにすぎないのです。この現象を自己免疫疾患として説明しようとする自己免疫疾患主義者の医者たちが世界中に存在していることは残念でたまりません。なぜ、自分たちの間違いに気が付かないのでしょうか???自己免疫疾患は絶対にありえないのです。
尿酸は、損傷したり、崩壊したりした細胞から放出される危険信号であるとも言えるのは、周辺の他の細胞にヘルペスが細胞に感染しているので自分の細胞にも感染する恐れがあることを知らせることができるからであります。核酸のプリン代謝物であるアデノシン三リン酸(ATP)と尿酸は、NLRファミリー(Nucleotide-bindingdomain,leucinerichrepeatcontainingreceptor)の1つである、NLRピリン-ドメイン含有蛋白質3(Nlrp3)インフラマソームを活性化してIL-1βとIL-18を誘導し、ヘルペス感染の危険信号となると同時に、アデノシン三リン酸(ATP)と尿酸の処理のための代謝経路とは別に大食細胞に貪食させてDAMPのゴミとして処理させるのです。DAMPとしてのアデノシン三リン酸(ATP)と尿酸は、自己免疫疾患の対象となるのではなくて、あくまでも屑としての単一の無駄な分子として大食細胞に処理されるだけなのです。しかも単一の分子であるアデノシン三リン酸(ATP)と尿酸は、自己免疫疾患の結果、壊れて細胞外に漏れ出た自己の成分では決してないのです。
タンパク質のDAMPの1つであるHMGB1とは何でしょうか?Highmobilitygroupboxprotein1の略語で、日本語で高移動グループボックスタンパク質1と訳します。高移動度群(HMG)ボックスタンパク質とは、細胞中のいたるところに豊富に存在する様々な機能を発揮するDNA結合タンパク質です。HMGB1はHMGタンパク質ファミリーのメンバーであり、典型的なクロマチン関連LSP(リーダーレス分泌タンパク質)です。LSP(leaderlesssecretoryprotein、訳してリーダーレス分泌タンパク質)とは、シグナルペプチドを持たないタンパク質で、DAMPの結果、潰れた細胞の外に放出されるので分泌されたように見えるDAMPのタンパク質の1つです。HMGB1は細菌によるエンドトキシンショックのために細胞が崩壊し、特定の免疫細胞によってDAMPとして認識され、大食細胞で処理されるために炎症反応を引き起こしているように見えます。TLR、TLR4、TLR9、およびRAGE(receptorforadvancedglycationendproducts、略してRAGE、訳して終末糖化産物受容体)に結合することによりNF-kB経路を活性化することにより炎症を誘発しているように見えます。
NF-kB経路とは何でしょうか? NF-κB(nuclearfactorkappa-light-chain-enhancerofactivatedBcells、訳して活性化B細胞核因子カッパ軽鎖転写促進因子、長すぎるので核内因子κBと訳される)は、DNAの転写、サイトカイン産生、細胞生存を制御するタンパク質複合体です。NF-κBはほとんどすべての動物細胞に見られ、ストレス、サイトカイン、フリーラジカル、重金属、紫外線照射、酸化LDL、細菌またはウイルス抗原などの刺激に対する細胞応答に関与しています。NF-κB(核内因子κB)は、感染に対する免疫応答の調節に重要な役割を果たしているのでNF-κBの作用を抑える薬は免疫を下げることになります。NF-κBは、シナプス可塑性と記憶のプロセスにも重要な働きを担っています。
NF-κBが活性化されるとRelBとヘテロ二量体化し、さらに遺伝子転写を促進するDNAと複合体を形成します。NF-κBは、まさに速効性の一次転写因子であり、不活性状態の細胞に存在し、活性化されるために新しいタンパク質合成を必要としない転写因子が即効性の細胞応答に役立っているのです。このような即効性の免疫反応を行うメンバーには、c-Jun、STAT、核内ホルモン受容体などの転写因子が含まれます。これにより、NF-κBは人体にとって有害な敵を最初に排除するための免疫反応を行うことができるのです。NF-κB活性を誘導する物質は非常に多様であり、活性酸素種(ROS)、腫瘍壊死因子アルファ(TNFα)、インターロイキン1-ベータ(IL-1β)、細菌性リポ多糖(LPS)、イソプロテレノール、コカイン、および電離放射線などの有害物質が含まれます。
刺激されていない細胞では、NF-κB二量体はIκB(κBの阻害剤)と呼ばれる阻害剤のファミリーによって細胞質に隔離されます。これは、アンキリンリピートと呼ばれる配列の複数のコピーを含むタンパク質です。
糖化とは何でしょうか? 糖化とは英語でGlycationといい、グルコース、フルクトースなどの糖がタンパク質または脂質への共有結合で生じる非酵素的なプロセスであります。糖尿病は糖化(Glycation)が起こりやすく、そのために糖尿病の三大合併症である「糖尿病網膜症」「糖尿病腎症」「糖尿病神経障害」が起こりやすく、いずれも動脈硬化による細小血管障害です。一方、大血管の糖化による動脈硬化は心筋梗塞や脳梗塞などを起こしやすいのです。糖化とは対照的に、グリコシル化は、タンパク質または脂質への糖の酵素媒介ATP依存性の結合です。グリコシル化は、タンパク質の翻訳後修飾の一般的な形態であり、成熟タンパク質の機能に必要です。グリコシル化にはN-結合型グリコシル化とO-結合型グリコシル化の2つのタイプが存在します。アスパラギン側鎖のアミドのN原子への付加はN-結合型グリコシル化、セリンとトレオニン側鎖のヒドロキシ基(OH)のO原子への付加はO-結合型グリコシル化によるものです。N-結合型グリコシル化とO-結合型グリコシル化についてはこちらを読んでください。DAMPのタンパク質のHMGB1とS100タンパク質について少し詳しく勉強しておきましょう。
まずHMGB1はCD80を増やすことによって樹状細胞の成熟を誘導できます。CD83、CD86、CD11c、および骨髄細胞における他の炎症誘発性サイトカイン(IL-1、TNF-a、IL-6、IL-8)の産生、および細胞接着分子(ICAM)や内皮細胞上の接着分子(VCAM-1)の発現増加もきたします。
次にS100タンパク質は、細胞内および細胞外の調節活動に関与するカルシウム調節タンパク質の多遺伝子ファミリーであり、癌による細胞の損傷や組織の損傷、特にヘルペスによる神経細胞の損傷の後にDAMPとして細胞内や細胞外に見られるのです。S100タンパク質の主な機能は、カルシウムの貯蔵とカルシウムをシャッフル(あちこちに動かす)ができます。それらは、細胞増殖、分化、移動、エネルギー代謝などのさまざまな機能を持っていますが、ヘルペスに感染した細胞が崩壊した後、それを貪食した大食細胞の受容体であるTLR2、TLR4、RAGEにS100タンパク質は認識されて、ヘルペスの感染に対する警戒危険信号として放出されたり、アミノ酸のゴミとしての放出されるDAMPタンパクの1つとなります。
ここで、PAMPやDAMPを認識するパターン認識受容体についてもう少し詳しく勉強しましょう。
パターン認識受容体、英語でPatternrecognitionreceptorで略してPRRは、自然免疫系の病原体が特有にもつ分子のパターンを認識して、人体に危険が侵入したというシグナルを自然免疫のみならず、獲得免疫(適応免疫)に伝える重要な機能を持っています。とりわけ現代の病原体は世界中の78億人が感染しているヘルペスが人体に存在していることを知らせるdangersignalです。
PRRは、病原体に典型的な分子を検出できる38億年かかって生殖細胞に記憶されて遺伝子に刻み込まれた(encode、訳してエンコード)感染宿主である人体のセンサー(hostsensor)です。PRR(Patternrecognitionreceptor)は主に、樹状細胞、マクロファージ、単球、好中球などの自然免疫細胞や、上皮細胞などによって発現されるタンパク質であり、病原体関連分子パターンと言われる病原体の持つPAMP(pathogen-associatedmolecularpattern、訳して、病原体関連分子パターン)とヘルペス感染細胞が崩壊した後のDAMP(damage-associatedmolecularpattern、訳してダメージ関連分子パターン)の2つのクラスの分子を認識するのです。
PAMP(pathogen-associatedmolecularpattern)は、微生物病原体、とりわけ現代は全ての人類に感染しているヘルペスウイルスの分子パターンを言い、これを認識するのがPRRであります。2つ目のDAMP(damage-associatedmolecularpattern)は、細胞の損傷または細胞死に至るまでに放出される宿主の細胞の成分が持つ、とりわけヘルペスが感染した細胞の成分の分子パターンであり、これをPRRで認識するのです。この自然免疫細胞の持つPRRは、適応免疫の前に進化したため、原始的なパターン認識受容体であります。大食細胞や樹状細胞などの抗原提示細胞(APC)が持つPRRは病原体に対しては病原体である敵と戦って殺すための炎症性サイトカインを放出し、自然免疫で殺しきれなければ抗原特異的な適応免疫応答につないでくれます。現代の最後に残る病原体は、99%がヘルペスウイルスであります。自己免疫疾患もヘルペスが原因です。自己の免疫は自己の成分を異物と認識できないので、自己免疫疾患は絶対にありえないのです。つまり、自己の成分は38億年かかって確立したPRRには認識されないので、自己免疫疾患はありえないのです。しかも、自己の成分はPAMPにも特別に異物として認識されるDAMPにも絶対になれないのです。
特定のPRRによって認識される微生物やヘルペスウイルスだけが持つ独特な特異的分子は、病原体関連分子パターン(PAMP)と呼ばれ、細菌の炭水化物(リポ多糖(LPS)やマンノース)と、核酸(細菌またはウイルスのDNAやRNA)と、フラジェリン(flagellin)と言われる細菌の鞭毛を構成するタンパク質の1種であるペプチドや、ペプチドグリカンとリポテイコ酸(グラム陽性菌由来)と、N-ホルミルメチオニンと、リポタンパク質および真菌グルカンとキチンなど数多くありますが自己の成分は含まれません。一方、内因性ストレス信号と言われたり、細胞障害性信号(屑)はダメージ関連分子パターン(DAMP)と呼ばれ、他の多くの化合物の中でも尿酸や細胞外ATPが含まれることは既に述べました。
PRRにはいくつかのサブグループがあり、リガンド特異性、機能、局在化、および進化的関係に従って分類されます。それらの局在に基づいて、PRRは、膜結合PRRおよび細胞質PRRに分類できます。膜結合型PRRには、1)Toll様受容体(TLR)と2)C型レクチン受容体(CLR)が含まれ、細胞質PRRには、3)NOD様受容体(Nod-like-receptor略してNLR)および4)RIG-I様受容体(RIG-I-like-receptor略してRLR)が含まれます。この4つのPRRについてもう一度詳しく復習しましょう。
1)Toll様受容体(Toll-like-receptor、略してTLR)として知られる膜貫通タンパク質によって媒介されて認識できるのは、細胞外またはエンドソームの病原体関連分子パターンです。TLRは現在知られているもので13種類あり、このTLRが最多のPRRです。TLRは、典型的な構造モチーフであるロイシンリッチリピート(LRR)を共有しており、特定の外観を持っています。TLR(Toll-like-receptor)は、サイトカインの合成と分泌を行い、病原体に対して自然免疫応答または獲得免疫応答の両方に必要な他の免疫防御のための細胞を活性化します。TLRファミリーの13種類の各々のTLRは、特定のPAMPをそれぞれ認識できます。
自己抗体は全てヘルペス感染細胞によって引き起こされたDAMP(細胞傷害関連分子パターン)を認識したPRR(パターン認識レセプター)によって生成されたmolecules(分子)によるものです。
受容体TLRや他のPRRによって認識される微生物と微生物成分と細胞の局在部位一覧表
| PRR(Pattern recognition receptor) | 局在部位 | 微生物成分 | 微生物 |
| TLR1/TLR2 | 細胞表面 | トリアシルリポペプチド | バクテリア |
| TLR2/TLR6 | 細胞表面 | ジアシルリポペプチド | マイコプラズマ |
| リポタイコ酸 | グラム陽性菌 | ||
| TLR2 | 細胞表面 | リポタンパク質 | さまざまな病原体 |
| ペプチドグリカン | グラム陽性菌と グラム陰性菌 | ||
| リポアラビノマンナン | マイコバクテリア | ||
| ポリン | ナイセリア | ||
| エンベロープ糖タンパク質 | ウイルス(例、はしかウイルス、HSV、サイトメガロウイルス) | ||
| GPI-ムチン | 原生動物 | ||
| ホスホリポマンナン | カンジダ | ||
| ザイモサン | 菌類 | ||
| β-グリカン | 菌類 | ||
| TLR3 | 細胞表面/ エンドソーム | dsRNA | ウイルス |
| TLR4 | 細胞表面 | LPS | グラム陰性菌 |
| エンベロープ糖タンパク質 | ウイルス(例、RSV) | ||
| グリコイノシトールリン脂質 | 原生動物 | ||
| マンナン | カンジダ | ||
| HSP70 | ホスト | ||
| TLR5 | 細胞表面 | フラジェリン | べん毛虫 |
| TLR7/TLR8 | エンドソーム | ssRNA | RNAウイルス |
| TLR9 | エンドソーム | CpGDNA | ウイルス、バクテリア、原生動物 |
| RIG-I | 細胞質 | dsRNA(短い)、5′-三リン酸RNA | ウイルス(例、A型インフルエンザウイルス、HCV、RSV) |
| MDA5 | 細胞質 | dsRNA(長い) | ウイルス(ピコルナウイルスおよびノロウイルス) |
| NOD1 | 細胞質 | ジアミノピメリン酸 | グラム陰性菌 |
| NOD2 | 細胞質 | MDP | グラム陽性菌とグラム陰性菌 |
| NALP1 | 細胞質 | MDP | グラム陽性菌とグラム陰性菌 |
| NALP3 | 細胞質 | ATP、尿酸結晶、RNA、DNA、MDP | ウイルス、バクテリア、およびホスト |
| DAI | 細胞質 | DNA | DNAウイルス、細胞内細菌 |
| AIM2 | 細胞質 | DNA | DNAウイルス |
| PKR | 細胞質 | dsRNA、5′-三リン酸RNA | ウイルス |
PRRの代表はTLRでありますが、上の一覧表には見慣れないTLR以外のPRRがあります。RIG-I、MDA5、NOD1、 NOD2、NALP1、NALP3、DAI、AIM2、PKRの9個のPRRについて順に説明しましょう。
1)RIG-Iはリグ-アイと読み、英語でretinoic acid-inducible gene-Iで、略してRIG-I(リグ-アイ)で、訳してレチノイン酸誘導遺伝子I(アイ)です。ヒトの自然免疫系で働くタンパク質の分子で、ウイルスが細胞内に進入した時にウイルス由来のRNAを認識し、抗ウイルス作用を示すI型インターフェロンであるIFN-αとIFN-βの産生を誘導します。自然免疫の中で細胞に感染したインフルエンザウイルスを細胞もろとも殺すことができるのはIFN-αとIFN-βだけであり、いかにRIG-Iの遺伝子の働きが極めて大事なのです。
2)MDA5、英語でMelanoma differentiation-associated gene 5で、略してMDA5で、訳してメラノーマ分化関連タンパク質遺伝子5です。様々な細胞内ウイルスセンサーであるMDA5タンパク質は、ウイルスが様々な細胞に感染した際にウイルスに由来する2本鎖RNAをこのPRRであるMDA5が認識して自然免疫応答を誘導するのです。MDA5はRIG-I様受容体(RIG-I like receptor、略してRLR)ファミリーの一部であり、RIG-IおよびLGP2も含まれており、ウイルスを検出できるパターン認識受容体として機能します。LGP2は、RIG-I様受容体3(RLR-3)としても知られ、RIG-IおよびMDA5によって認識される多くのウイルスに対して効果的な抗ウイルス応答を生成するために不可欠なのです。つまり、RIG-Iが作るIFN-αとIFN-βを作るためにはLGP2の手助けが絶対に必要なのです。
RIG-IとMDA5は、互いに相同性の高いヘリカーゼドメインを有していますが、認識できるRNAの構造や配列(pathogen-associated molecular pattern、PAMP)が異なっており、HCV(C型肝炎ウイルス)はRIG-IとMDA5いずれにも認識されます。ヘリカーゼドメインは、一本の鎖でできているRNAが入り組んだ構造をとり、部分的に結合して2本の鎖になっている構造のRNAの部分を紐解いて、本来の一本の鎖に戻す働きを持つタンパク酵素をヘリカーゼと呼びます。ヘリカーゼの活性のある中心部分をヘリカーゼドメインといいます。
I型インターフェロンのかかわる自然免疫応答は,外部から侵入したヘルペスウイルスなどの微生物を自然免疫受容体により感知し,その排除に働く一方、細胞の表面やエンドソームにはToll様受容体(Toll-likereceptor、略してTLR)が存在し,病原体関連分子パターン(pathogen-associated molecular pattern、PAMP)を認識することにより抗ウイルス応答、抗ヘルペスウイルス応答を起こして、細胞もろともヘルペスウイルスを殺してしまうのです。
3)NOD1は英語でNucleotide-binding oligomerization domain-containing protein 1で、略してNOD1で、訳してヌクレオチド結合オリゴマー化ドメイン含有タンパク質1です。oligomerization は何でしょうか? 日本語もオリゴメライゼーションで、oligomerization によって生ずるオリゴマー化は、有限の重合度(ポリマー化)によってモノマー(単量体)を高分子複合体に変換させる化学プロセスです。簡単に言えば、オリゴメライゼーションはモノマー(単量体)をオリゴマー化することです。
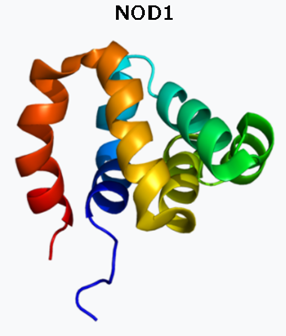
ヌクレオチド結合オリゴマー化ドメイン含有タンパク質1(NOD1)は、ヒトではNOD1遺伝子によってコードされるタンパク質受容体です。細胞内のバクテリアの分子をPRRで認識し、バクテリアを殺すための免疫反応を刺激するのです。NOD1タンパク質には、カスパーゼ動員ドメイン(CARD)が含まれています。カスパーゼ動員ドメイン(CARD)とは何でしょうか? CARDは英語でCaspase recruitment domainsで、さまざまなタンパク質、炎症およびアポトーシスに関連するプロセスに関与するタンパク質に見られる相互作用モチーフです。カスパーゼ(Caspase)は、システイン-アスパラギン酸プロテアーゼや、システイン依存性アスパラギン酸指向性プロテアーゼとも言われ、プログラムされた細胞死に不可欠な役割を果たすプロテアーゼ酵素のファミリーです。個々のCARD間の直接的な相互作用を介して、より大きなタンパク質複合体の形成を仲介します。CARDドメインは、ヘリカーゼ、キナーゼ、ミトコンドリアタンパク質、カスパーゼ、その他の細胞質因子など、驚くほど広範囲のタンパク質に見られます。
NOD1はNOD様受容体タンパク質ファミリーのメンバーであり、次に説明するNOD2の親戚と言えます。NOD1は細胞内パターン認識受容体であり、植物の耐性タンパク質と構造が類似しており、細菌ペプチドグリカンを含む細菌のD-グルタミル-メソ-ジアミノピメリン酸(iE-DAP)分子を認識することにより、自然免疫と獲得免疫を仲介します。植物の耐性タンパク質とは、植物は、生育に適した環境では最大限に生長する一方、周辺環境が悪化すると生長を抑え、環境ストレスに耐えるためのタンパク質を持っており、これを耐性タンパク質といいます。
NOD1はRIPK2と相互作用します両方の分子のCARDを介して。iE-DAPを含む分子によるNOD1の刺激は、転写因子NF-κBの活性化をもたらします。RIPK2とは、英語でReceptor-interacting serine/ threonine-protein kinase 2で、日本語で分かりやすく訳すと「受容体と相互作用するセリンもしくはスレオニン-プロテインキナーゼ2です。ヒトではRIPK2遺伝子によってコードされる酵素です。酵素キナーゼタンパク質は、C末端カスパーゼ動員ドメイン(CARD)を含み、自然免疫経路と適応免疫経路の両方におけるシグナル伝達複合体の構成要素で、自然免疫と獲得免疫を仲介します。NF-κBの強力な活性化因子であり、さまざまな刺激に応答してアポトーシスを誘導します。
NF-κBとは何でしょうか? NF-κBは英語でnuclear factor-kappa Bで、訳して核内因子κBで、読み方はエヌエフ・カッパー・ビーです。NF-κBは1986年に、David BaltimoreらによりB細胞で選択的に発現する免疫グロブリンのκ軽鎖のエンハンサーに結合する転写因子として同定されました。DNA上の塩基配列領域にあるエンハンサーは遺伝子の発現制御において中心的な役割を担っており、このエンハンサーに転写因子が結合すると、標的遺伝子の発現にスイッチが入り、タンパク質を生成するのです。エンハンサーは、遺伝子活性化因子である転写因子と結合することで遺伝子の転写量を大幅に増大(enhance)させることから、エンハンサーと命名されました。
発見当初はB細胞に特異的なものだと考えられていたのでNFにB細胞のBと、B細胞の軽鎖の1つであるκ(カッパー)が付け加えられたのです。ところが後にB細胞のみならず動物のほとんど全ての細胞に発現していることがわかりました。しかも高等生物に限らずショウジョウバエやウニなどの無脊椎動物の細胞においてもNF-κBが発現しています。NF-κBはストレスやサイトカイン、紫外線等の刺激により活性化されます。言うまでもなく、NF-κBは免疫反応において中心的役割を果たす転写因子の一つであり、急性および慢性炎症反応や細胞増殖、アポトーシスなどの数多くの生理現象に関与しています。転写因子というのはタンパク質の設計図である遺伝子であるDNAやRNAをタンパク質の合成の指示書であるmRNAの遺伝子に移し替える(転写する)ことです。NF-κB活性制御の不良はクローン病や関節リウマチなどの炎症性疾患をはじめとし、癌や敗血症性ショックなどの原因となり、特に悪性腫瘍では多くの場合NF-κBの恒常的活性化が認められると言われていますが、これらの病気にはとりわけヘルペスウイルスが絡んでおり、その結果、免疫の要であるNF-κB活性制御の不良が起こるからです。人類最後に残る病原体はヘルペスウイルスであり、ヘルペスウイルスを増殖させるようなNF-κB活性制御の不良は免疫の低下によってもたらされ、免疫の低下が改善される時に増殖したヘルペスウイルスとの戦いが始まることによってあらゆる原因不明の病気やありえない自己免疫疾患が増え続けることになるのです。NF-κBの働きが人為的に抑制されてヘルペスウイルスによる自己免疫疾患がどのように起こるかについては別のタイトルで説明しましょう。
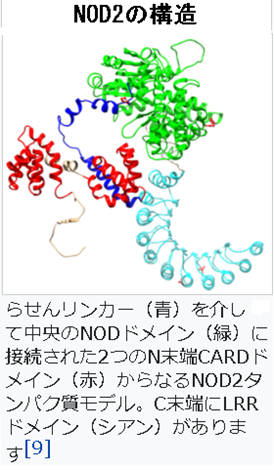
4)NOD2は、英語で Nucleotide-binding oligomerization domain-containing protein 2 (NOD2)カスパーゼ動員ドメイン含有タンパク質15(CARD15)または炎症性腸疾患タンパク質1(IBD1)としても知られるヌクレオチド結合オリゴマー化ドメイン含有タンパク質2(NOD2)は、細胞内のバクテリアの分子のペプチドグリカンを認識するPRRであり、認識したのちに免疫反応を刺激する重要な役割を果たしています。NOD2は細胞内パターン認識受容体であり、植物の耐性タンパク質と構造が類似しており、特定の細菌に見られるムラミルジペプチド(MDP)と呼ばれる特定の構造を含む分子を認識します。ムラミルジペプチド(Muramyl di-peptide、略してMDP)は、グラム陽性菌とグラム陰性菌の両方に存在するペプチドグリカンの一つです。
タンパク質のC末端部分には、タンパク質間相互作用で役割を果たすロイシンリッチリピートドメインが含まれています。NOD2のタンパク質の中央部分は、タンパク質の自己オリゴマー化に関与する特徴的なNODドメインを持っています。N末端部分は、アポトーシスおよびNF-κB活性化経路において役割を果たす2つのCARDドメインが含まれています。上左図にNOD2の構造を示しておきます。ここでのリンカーというのは、2つのドメインを結びつけているタンパク質です。
このNOD2の遺伝子は、NOD1 / Apaf-1ファミリーと知られているNOD様受容体ファミリーで、2つのカスパーゼ動員ドメイン(CARD)と11のロイシンリッチリピート(LRR)を持つタンパク質をコードします。このタンパク質は主に末梢血白血球で発現し、グラム陽性菌とグラム陰性菌の両方に存在するムラミルジペプチド(MDP)部分を有する細菌分子を認識し、NF-κBタンパク質を活性化することにより、免疫応答に関与します。この遺伝子の変異は、関連付けられているクローン病、ブラウ症候群、重度の肺サルコイドーシス 及び移植片対宿主病に関与しています。ブラウ症候群は、皮膚と関節そして眼にサルコイドーシスと呼ばれる病気によく似た 肉芽腫 ができる病気です。
NOD2はNLRC4と相互作用することが示されています。NOD2は、ssRNAまたはウイルスRNA処理に応答してMAVSに結合し、ヘルペスウイルスに対してIFN-αやIFN-β産生を活性化できます。

5)NALP1は英語でNACHT leucine-rich-repeat protein1で、略してNALP1です。HACHTとは、その名前の由来は、NAIP(NLPファミリーアポトーシス阻害タンパク質)のNAと、CIITA(つまり、C2TAまたはMHCクラスII転写活性化因子)のCと、HET-E(カビの一種である糸状菌のPodospora anserinaから見つけられた非互換性遺伝子座タンパク質)のHと、最後のTはTEP1(TP1またはテロメラーゼ関連タンパク質)のTから生まれました。NLRP1は、主に上皮細胞や造血細胞に発現していますが、他に、小腸、胃、気道上皮の腺上皮構造内に見られます。NLRP1の遺伝子は、プログラム細胞死であるアポトーシスに関与しています。NLRP1は、抗菌または抗ヘルペスウイルス免疫応答を活性化します。
6)NALP3は英語でNACHT leucine-rich-repeat protein3で、NLRP3は、マクロファージで主に発現され、インフラソームの成分として見られます。インフラソームとは、英語でinflammasomeで、主に自然免疫細胞を中心に発現し、複数のタンパク質からなるタンパク複合体です。病原微生物成分や尿酸結晶などの細胞質内の異物をすでに説明したNOD-like receptors(NLRs)に分類されるNLRC4(NLR Family CARD Domain Containing 4)、NLRP1(NLR Family Pyrin Domain Containing 1)、NLRP3(NLR Family Pyrin Domain Containing 3)などを介して宿主細胞に対するdanger signal(危険信号)として認識するPRRであり、シグナル伝達分子apoptosis-associated speck-like protein containing a CARD(下線の頭文字を採用して略してASC)を介して、非活性型のprocaspase-1を活性型のcaspase-1にし、casepase-1はpro-IL-1βやpro- IL-18を炎症性サイトカインとして実際に働くIL-1βやpro- IL-18にし、炎症反応の誘導や進展に重要な役割を果たしています。NLRP3は細胞外のATPや結晶化した尿酸のような、傷害された細胞の産物を見つけ出すことができます。
NLRP3の遺伝子は、ピリン(pyrin)ドメインやヌクレオチド結合部位(nucleotide-binding-site、略してNBS)ドメインやロイシンリッチリピート(Leucine-rich repeat、略してLRR)モチーフの3つを含んでいるピリン様プロテイン(Pyrin like protein)をコードしています。このNLRP3タンパク質は、ASCと言われるapoptosis-associated speck-like protein containing a CARD、日本語で「CARDを含むアポトーシス関連スペック様タンパク質」と訳すのですが、このASCというタンパク質とピリンドメイン(Pyrin domain、略してPYD)は相互作用します。カスパーゼ動員ドメインであるCARD(caspase recruitment domain、訳してカスパーゼ誘導ドメイン)を含むタンパク質は、炎症と免疫応答とアポトーシスとに関与します。ピリンドメイン(PYD)とは、ピリンドメインがあるタンパク質ドメインと、アポトーシスに関わる三次元の立体構造であるdeath domain(死のドメイン)と言われるサブクラスタンパク質モチーフから成り立っています。ピリンドメインを持つタンパク質には、NOD様受容体と呼ばれる細胞内微生物センサーや、ASC(apoptosis-associated speck-like protein containing a CARD)の機能に関連するタンパク質が含まれます。
NLRP3(NOD-,LRR- and pyrin domain-containing protein 3)は、病原体関連分子パターン(PAMP)を認識するパターン認識受容体(PRR)として機能する自然免疫系の成分です。NLRP3はPRRであるNOD様受容体(NOD like receptor、NLR)サブファミリーに属しています。NLRP3 はアダプターASCタンパク質であるPYCARDと共にカスパーゼ-1活性化複合体を形成します。この複合体はNLRP3インフラソームと呼ばれます。アダプタータンパク質とは何でしょうか? アダプタータンパク質、英語でAdaptor ProteinとかAdaptor Molecule といい、シグナル伝達に関与するタンパク質です。アダプタータンパク質自体は基本的に酵素活性を有していないが、他のタンパク質との結合に関与するドメインを複数有しており、SH2ドメインやロイシンジッパー、Znフィンガー、ASCタンパクなどがあります。アダプタータンパク質はこれらの構造を介してシグナル伝達分子と結合し、そのリクルート(招集)を行うと共に、受容体とシグナル伝達分子の会合を仲介する役割を持ち、チロシンキナーゼや酵素などのシグナル伝達分子を受容体の近くに集めることでシグナル伝達分子の活性化を促進させるのです。インフラソームとは何でしょうか? 炎症は生体に傷害をもたらす病原体に対する防御の戦いの反応です。この生体防御反応に対して炎症性サイトカインが作られ、とくに重要な炎症性サイトカインの1つがインターロイキン1β(IL-1β)であり、このIL-1β産生を制御する細胞内のタンパク質複合体をインフラソームと言います。それではIL-1βとは何でしょうか? インターロイキン1ベータ(IL-1β)は、1)白血球パイロジェン、2)白血球内因性メディエーター、3)単核細胞因子、4)リンパ球活性化因子などの他の4つの名前で知られるサイトカインタンパク質です。このIL-1βサイトカインは、活性化されたマクロファージによってプロタンパク質(protein precursor、訳してタンパク前駆物質)として産生され、カスパーゼ1(CASP1、別名Interleukin-1 converting enzyme、略してICE)によって活性型にプロセシングされます。このIL-1βサイトカインは炎症反応の重要なメディエーターであり、細胞増殖、分化、アポトーシスなどのさまざまな細胞活動に関与しています。中枢神経系(Central nervous system、略してCNS)におけるこのIL-1βサイトカインによるシクロオキシゲナーゼ-2(cyclooxygenase 2、略して COX2と別に Prostaglandin-endoperoxide G synthase 2 、略して PTGS2とも言います。)の誘導は、炎症性疼痛過敏症に寄与することは分かっていますが、疼痛そのものを起こすわけではありません。インターロイキン-1(IL-1)ファミリーサイトカインは、11種の遺伝子 によってコードされる11種のタンパク質(IL-1Family1~IL-1Family11)から成り立ち、炎症性サイトカインの複雑なネットワークを誘導し、白血球と内皮細胞でのインテグリンの発現を介して、炎症反応を調節および開始します。インテグリンとは、細胞間および細胞外マトリックスの接着を促進する膜貫通型受容体です。
さまざまなインフラソーム複合体(細胞質ゾル分子複合体)が存在し、それぞれのインフラソームは危険信号を認識し、炎症誘発性プロセスとIL-1βおよびIL-18の産生を活性化します。 NLRP3(ピリンドメイン、ヌクレオチド結合ドメイン、ロイシンリッチリピートの3つのドメインを含んでいます。)タイプのインフラマソームはさまざまな刺激によって活性化され、2型糖尿病、アルツハイマー病、肥満、アテローム性動脈硬化症など、NLRP3の活性化に関連する疾患があります。しかしながら、NLRP3(NOD-like receptor )もDAMPを認識するPRRの1つですからこれらの病気の原因にはならないのです。
IL-1αとは何でしょうか? ヘマトポエチン1としても知られるインターロイキン1アルファ(IL-1α)は、ヒトではIL1A遺伝子によってコードされる11種類あるインターロイキン1ファミリーのサイトカインの1つです。一般に、インターロイキン1は、炎症の発生、および発熱と敗血症の促進に関与しています。IL-1α阻害剤は、これらのプロセスを中断し、病気を治療するために開発されています。しかしこのようなIL-1αは病気の原因になるのではなくて IL-1αは、IL-1α阻害剤を治療薬に用いても根本的な治療にはならないのです。なぜならば全ての免疫のサイトカインは病原体を処理するために作用しているだけですから、どんな病原体がIL-1αを活性化しているかをまず考える必要があります。現代社会において最後に残る病原体はヘルペスウイルスであることを知っておく必要があります。だからこそIL-1αはヘルペスウイルスを処理するために、主に活性化マクロファージ、好中球、上皮細胞、血管内皮細胞によって産生されるのです。それは、代謝的、生理学的、造血活性を有し、免疫応答の調節において中心的な役割の1つを果たしています。インターロイキン-1受容体(Interleukin-1 receptor、略してIL-1R)に結合します。腫瘍壊死因子-αを活性化する経路上にIL-1Rが存在しています。
インフラマソームは、カスパーゼ1の活性化を調節し、ヘルペスウイルスなどの感染性微生物やDAMPの産物として宿主タンパク質に由来する分子に応答して炎症のクズを処理するための自然免疫系の受容体およびセンサーがインフラソームであって、決して炎症性疾患を起こすものではなくて掃除屋であるということはすでに述べました。
外敵との戦いがない時には、活性化シグナルが生まれない場合にはNLRP3は、細胞質内でHSP90はSGT1と複合体を形成した不活性状態に保たれています。HSP90とは、Hsp90(熱ショックタンパク質90)は、他のタンパク質が適切に折りたたまれるのを助け、熱ストレスに対してタンパク質を安定させ、タンパク質分解を助けるシャペロンタンパク質です。シャペロンタンパク質とは、すべての哺乳類タンパク質の半分以上の折り畳み(fold)に関与しており、高分子構造の組み立てまたは分解を支援するタンパク質です。SGT1とは、英語でsuppressor of G-two allele of Skp1、略してSGT1で、複数の生物学的機能を持つアダプタータンパク質です。SGT1はNod1の活性化に不可欠です。
NLRP3インフラマソームは傷害した細胞などから放出された結晶尿酸と細胞外ATPなどを異物信号として検出します。これらのシグナルは、インフラソーム複合体からHSP90とSGT1を放出させ、かつインフラソーム複合体へASCタンパク質とカスパーゼ-1を動員します。活性化されたNLRP3インフラソーム複合体の中にあるカスパーゼ-1は、炎症性サイトカインであるIL-1βを活性化し、炎症後のゴミの処理をします。
7)DAIは、英語でDNA-dependent activator of IFN-regulatory factors、略してDAI、訳してDNA依存性IFN調節因子のアクチベーターです。DAIはZ-DNA結合タンパク質1(Z-DNA-binding protein 1、略してZBP1)やIFN調節因子やDLM-1やDNA依存性活性化因子としても知られており、ヒトではZBP1遺伝子によってコードされるタンパク質です。したがって、DAIにはトータルで5つの名称があります。別の4つの名称は、1つ目がZ-DNA結合タンパク質1(ZBP1)、2つ目がIFN調節因子、3つ目がDLM-1、4つ目がDNA依存性活性化因子となり、DAIを含めて5つの名称があります。
DAIは、細胞質内のウイルスDNAを認識します。ウイルスのライフサイクルには、多くの場合、ウイルスのDNAやRNAが細胞質に露出するステップが含まれますが、細胞のDNAは通常、細胞の核に含まれているため、細胞はウイルス感染の指標としてDAIなどのタンパク質を利用して、ウイルスの遺伝子を認識するのです。DAIが活性化されると、インターフェロンβ(IFN-β)やインターフェロンα(IFN-α)などの抗ウイルス性サイトカインの産生が増えます。細胞内のヘルペスウイルス感染やA型インフルエンザウイルス感染を感知し、細胞死をもたらします。この細胞死が後で説明するようにネクロプトーシスとかネクロトーシスとか言われるのです。
ネクロトーシスやネクロプトーシスとは一体何でしょうか? ZBP1(DAI)がA型インフルエンザウイルス(IAV)感染中に生成されたZ型RNAを感知し、ネクロトーシスと呼ばれるカスパーゼ非依存性の炎症性細胞死が起こることが最近、発見されました。ZBP1(DAI)がA型インフルエンザウイルス(IAV)のリボ核タンパク質複合体を感知して細胞死を誘導するのです。リボ核タンパク質とは、リボ核酸タンパク質ともいい、リボ核酸というのは英語でribonucleic acidといい、RNAのことです。このリボ核酸(RNA)とタンパク質が結合した複合体をリボヌクレオタンパク質、英語でRibo-nucleo-proteinといい、略してRNPと呼ばれるのです。転写因子であるIRF1(Interferon regulatory factor 1、訳してインターフェロン制御因子1)がZBP1(DAI)発現の上流調節因子である転写因子です。
Z型RNAとは何でしょうか? ちょうどDNAの二重らせん構造の取り方に3種類のA-DNA、B-DNA、Z-DNAがあるように、RNAにもDNAと同様にらせん構造があり、その構造の取り方の1つにZ-RNAがあるのです。リボ核酸は英語でribonucleic acidで、略してRNAは、単一の(モノマー)のリボヌクレオチド(RNA)が何個もホスホジエステル結合で重合して鎖状に繋がった核酸です。ちょうどDNAは2個のDNAのヌクレオチドをリン酸がホスホジエステル結合で結びつけることを繰り返して何個も結びつけて鎖状にしたのがDNAなのです。1個のRNAヌクレオチドはリボース、リン酸、塩基の3つの成分から構成され、基本的に核酸塩基としてアデニン (A)、グアニン (G)、シトシン (C)、ウラシル (U) を有していますが、DNAはウラシル (U) がチミン(T)になっている違いがあるだけです。RNAポリメラーゼによりDNAを鋳型にして転写(合成)されてmRNAになります。RNAは生体内でタンパク質合成を行う際に必要なリボソームの活性中心部位を構成しています。現在、新型コロナウイルスワクチンのmRNAはリボゾームで新型コロナウイルスのスパイクを作らせ、このウイルスのスパイクのタンパク質に対して人間の免疫がIgM抗体を作っているのですが、IgG抗体はmRNAワクチンでは作れないのでせいぜいワクチンとしての効果が続くのは6か月以内でIgMが消えてしまうので本当のワクチンではないのです。
RNAには色々種類があり、生体内での挙動や構造により、RNAには、伝令RNA(メッセンジャーRNA、mRNA)、運搬RNA(トランスファーRNA、tRNA)、リボソームRNA (rRNA)、ノンコーディングRNA (ncRNA)、リボザイム、二重鎖RNA(dsRNA) 等の様々な分類があります。
DNAのらせん構造の3種類のA-DNA、B-DNA、Z-DNAについて説明しましょう。 A-DNAは、B-DNAに似た右巻き二重らせんですが、B-DNAよりも短くコンパクトならせん構造であり、塩基対はらせん軸に対して直交していません。A型とB型のDNA構造は本当はワトソンとクリックと一緒にノーベル賞をもらうべきであったロザリンド・フランクリンによって発見されました。彼女こそがDNAは二重らせん構造が遺伝子であることを証明したのです。さらにロザリンド・フランクリンは脱水条件下でA型のDNA構造となることも示しました。こうした条件はDNAの結晶の形成の際によく利用され、多くのDNAの結晶構造がA型構造です。同様のらせん構造は、RNAの二重らせんやDNA-RNAハイブリッドの二重らせんでもみられるのです。
ネクロトーシスとは、プログラムされた形態の壊死、または炎症性細胞死です。従来、壊死は、アポトーシスを介した秩序だったプログラム細胞死とは対照的に、細胞損傷または病原体による浸潤破壊によって起こるプログラムされていない細胞死です。ネクロトーシスの発見は、細胞がプログラムされた方法で壊死を実行できるのみならず、アポトーシスが必ずしも細胞死の好ましい形態であるとは限らないのです。さらに、ネクロトーシスの免疫原性(免疫を活性化する性質)は、免疫系による病原体に対する防御を助けるなど、特定の状況でのネクロトーシスの関与に有利に働きます。なぜならば、ネクロトーシスによって感染した細胞はウイルス(ヘルペス)と共にネクロトーシスしてしまうのですが、新しい細胞に感染させないので生体細胞に対する感染拡大を防ぐからです。ネクロトーシスはヘルペスウイルス防御メカニズムとして明確に定義されており、ウイルス複製を制限するウイルスカスパーゼ阻害剤の存在下で、細胞がカスパーゼ非依存的に「細胞自殺」を起こすことが可能なのがネクロトーシスなのです。ネクロトーシスは、疾患への反応であることに加えて、クローン病、膵炎、心筋梗塞などの炎症性疾患の構成要素としても特徴づけられています。つまりこのような病気にはヘルペスが絡んでいることを示唆しているのです。言い換えると、クローン病のような腹痛をもたらす自己免疫疾患もヘルペスが関与していると考えるべきです。このようなネクロトーシスの代表はヘルペス感染細胞が自らIFN-αやIFN-βを産生し、自らの細胞もろともヘルペスを博愛主義的な細胞自殺に追い込むカスパーゼ非依存的な殺し方と言えます。
ヘルペスウイルスがネクロトーシスをもたらすシグナル伝達経路について説明しましょう。 その経路はまず体内にヘルペスウイルスが感染し、大食細胞に貪食され、大食細胞のToll like receptorに認識されるとTNFαが作られ、血中に放出されるとあらゆる細胞の細胞膜に存在するその受容体TNFR1(Tumor necrosis factor receptor 1)を刺激します。TNFR1(Tumor necrosis factor receptor 1)結合タンパク質TNFR関連死タンパク質であるTRADD(Tumor necrosis factor receptor type 1-associated DEATH domain)およびTNF受容体関連因子TRAF2(TNF receptor-associated factor 2)はRIPK1(Receptor-interacting serine/threonine-protein kinase 1)にシグナルを送り、RIPK3(Receptor-interacting serine/threonine-protein kinase 3)を動員してネクロソームを形成します。ネクロソームはリポプトソームとも呼ばれます。ネクロソームとは、ネクローシスを誘導するシグナル伝達複合体であります。ネクロソーム複合体に含まれているタンパク質にはRIPK1(receptor-interacting protein kinase 1)と, RIPK3(receptor-interacting protein kinase 3)と, MLKL(mixed lineage kinase domain-like protein)です。リポプトソームは、ネクロプトーシスとアポトーシスによる細胞死を活性化するのです。リポプトソームによるMLKL(mixed lineage kinase domain-like protein)のリン酸化は、MLKLのオリゴマー化(MLKLがいくか繋がること)を促進し、MLKLが細胞の原形質膜および細胞小器官に挿入され、MLKLが自由に透過できるようになってしまい、MLKLが細胞の原形質膜および細胞小器官と一体化することにより、炎症を起こしている状態となり、様々のダメージ関連分子パターン(DAMP)が放出され、後始末のための免疫応答が誘発されます。免役応答には敵との戦いによって正常な細胞が崩壊したあとの屑であるDAMPも処理する仕事も含まれているのです。
8)AIM2、英語でabsent in melanoma 2で、略してAIM2です。AIM2は細胞質内で二本鎖DNAを認識する受容体であり,PRRの1つであり、カスパーゼ1の活性化を介して戦いの後の処理をする炎症性サイトカインの産生や炎症性プログラム細胞死を誘導します。
IFN-αやIFN-βなどのインターフェロン誘導性タンパク質であるAIM2は、ヒトではAIM2遺伝子によってコードされます。AIM2は、インフラマソームの一部であり、細菌のDNAおよびヘルペスウイルスのDNAに対する防御に貢献しています。AIM2は、アミノ酸1〜87個から成り立っているN末端のDAPIN(またはピリン)ドメインとアミノ酸138〜337から成り立っているC末端のHIN-200ドメインを持つ343個のアミノ酸から成り立っているタンパク質で、2つのオリゴヌクレオチドと結合してる皺襞(ひだ)を持っているのがAIM2というPRR(Pattern recognition receptor、訳してパターン認識受容体)です。病原体や腫瘍を認識すると、その病原体や腫瘍を駆逐するために様々な機能を発揮するのです。
AIM2は、IFN-γ(インターフェロン-ガンマ)によって発現が誘導され、腫瘍形成を起こす細胞増殖を制御するのです。インターフェロンガンマはAIM2の発現を誘導します。タンパク質AIM2のC末端のHINドメインは、ヘルペスウイルスや細菌や宿主の二本鎖DNAに結合し、細胞質ゾルのdsDNAセンサーとしてdanger signalとして機能します。これにより、インフラマソーム複合体がオリゴマー化されます。AIM2のN末端ピリンドメインは、別のタンパク質ASC(Apoptosis-associated speck-like protein containing a CARD、訳してCARDドメインを含んでいるアポトーシス関連スペック様タンパク質)のピリンドメインと相互作用します。ASCには、プロカスパーゼ-1を複合体まで動員するCARDドメイン(カスパーゼ活性化ドメインと動員ドメイン)を含んでいます。ASCのこの働きにより、カスパーゼ-1が自動活性化されます。このカスパーゼ1は炎症性サイトカインであるIL-1bおよびIL-18を加工する酵素です。核膜の正常な状態が薬理学的破壊によって。AIM2インフラマソームは活性化されます。というのは、核膜が破壊されると核内の様々な成分がDAMPとしてAIM2に認識されるからインフラソームが活性化されるのです。AIM2発現レベルの上昇は、ヘルペスによって生ずる乾癬患者の皮膚細胞に見られます。乾癬(かんせん)とは、ヘルペス感染細胞がヘルペスに破壊されると皮膚から少し盛り上がった赤い発疹(ほっしん)の上に、銀白色のフケのような垢(あか)が付着し、ポロポロとはがれ落ちる病気です。全身性エリテマトーデス(SLE)において見られるヘルペスによるリソソーム機能不全は、核内から漏れ出たDNAが細胞質ゾルへ漏れ出て、AIM2を活性化させてしまい、その結果1型のインターフェロン(IFN-α、IFN-β)が増えてしまうのです。増えた1型のインターフェロンはヘルペスが隣接する細胞に感染しないようにPRRであるAIM2が働きだすのです。リソソームとは、酸性に保たれた細胞内小器官(オルガネラ)の一つであり,複合糖質や脂質などの細胞内基質の分解に関わり、リソソーム内部には酸性pH条件下で働く種々の加水分解酵素(リソソーム酵素)が存在し,エンドサイトーシスやオートファジーにより細胞内外から取り込まれた生体分子を加水分解します。分解されて生成したアミノ酸,糖そして脂質は再利用されるので、リソソームは細胞内の「リサイクル工場」なのです。しかし、ヘルペスによってリソソームも破壊されると、リソソーム機能不全が生じるのです。
全身性エリテマトーデス(SLE)は、自己免疫疾患の代表ですが、そもそも自己免疫疾患は絶対にありえない病気です。それではどのようにして自己免疫疾患の証拠である抗核抗体が生まれるのでしょうか? ヘルペスによって破壊された核内から漏れ出たDAMPの一分子である核の成分をたまたま認識したBリンパ球がヘルパーT細胞に依存せず認識し作りだして血中に放出した分泌IgMクラスの抗核抗体がSLEの患者に見られた時、自己免疫疾患の証拠と言い張っているのが現代の自己免疫論者の間違った主張なのです。あくまでもヘルペスによって核が破壊されたDAMPの分子にすぎない核の成分が全ての自己免疫疾患の原因であることに気がつかないのです。言い換えると、原因と結果を逆にしてしまっているのです。例えばアルツハイマーの病気の原因もヘルペスが海馬の細胞に感染した時にヘルペスの増殖と共に傷ついたり破壊された後に細胞から漏れ出た残骸、つまりDAMPがアミロイドβであるのにアミロイドβ自身がアルツハイマーの原因であると言っているのと同じなのです。
このように自己免疫疾患や原因不明の病気は全てヘルペスによるものであることを認めない限り永遠に現代の病気を治す切ることは不可能でしょう。病気を治すことほど簡単なことはないのに。残念です。
9)PKR(Protein kinase R) プロテインキナーゼR(PKR)は、セリン-スレオニンキナーゼであり、 mRNAの翻訳、転写制御、アポトーシスの調節、および細胞増殖のような主要な役割を果たしています。そのようなPKRの働きの調節不全は、癌や、神経変性、炎症、および代謝障害に関わっています。
PKRはインターフェロンを誘導することによって、ウイルス感染から細胞を保護します。
プロテインキナーゼR(Protein kinase R、略してPKR)は、ウイルス感染によって細胞に導入されたウイルスの二本鎖RNA(dsRNA)によって活性化されます。PKRには、N末端のdsRNA結合ドメイン(Double-stranded RNA-binding domain、略してdsRBD、訳して二重鎖のRNAに結合するドメイン)とC末端の キナーゼドメインが含まれており、アポトーシス促進(計画された細胞死)機能を持っています。dsRBD(Double-stranded RNA-binding domain)は、保存された二本鎖RNA結合モチーフの2つのタンデムコピー(tandem copy)であるdsRBM1とdsRBM2の2つのモチーフで構成されています。dsRBM1は英語でdouble-stranded RNA-binding motif 1であり、dsRBM2はdouble-stranded RNA-binding motif 2です。タンデムコピー(tandem copy)とは、dsRBM1とdsRBM2の2つのモチーフの末端に、突出末端部の配列が同じ酵素部位を付加し2個のモチーフが結合して縦列に並ぶことによって、コピー同士が切り離せないフラグメントになることです。
ヘルペスなどのウイルス感染の状況では、ウイルス複製と遺伝子発現によって作成されたdsRNAがN末端ドメインに結合し、タンパク質を活性化します。活性化すると、PKRは真核生物の翻訳開始因子eIF2α(Eukaryotic Initiation Factor 2)をリン酸化することができます。これはさらなる細胞のmRNA翻訳を阻害し、それによってウイルスタンパク質合成を防ぎます。ElF2α(Eukaryotic Initiation Factor 2、略してeIF2)は複製開始コドンであるAUGコドン(メチオニン)からの一般的な開始翻訳に関与しているため、代わりに代替の非AUG開始が行われます。非AUG開始を使用するmRNAの例は、熱ショックタンパク質のmRNAです。活性化PKRは、その阻害サブユニットであるIkBをリン酸化することにより、転写因子NFkBの活性化を仲介することもできます。活性化されたNFkBは、インターフェロンサイトカインの発現をアップレギュレートします。インターフェロンサイトカインは、抗ウイルスシグナルを局所的に拡散する働きをします。活性化PKRは、細胞周期と代謝を調節する腫瘍抑制因子PP2Aを活性化することもできます。複雑なメカニズムを通じて、活性化PKRは細胞アポトーシスを誘発し、さらなるウイルスの拡散を防ぐこともできます。 PKRストレス経路の中で、PKRは、病原体、栄養素の不足、サイトカイン、照射、機械的ストレス、小胞体ストレスなどのさまざまなストレスシグナルに対する細胞応答の中心にあります。PKR経路は、JNK、p38、NFkB、PP2A、eIF2αのリン酸化などの他のストレス経路の活性化を通じてストレス応答を引き起こします。折りたたまれていないタンパク質の過剰によって引き起こされる小胞体ストレスは、炎症反応を引き起こします。PKRは、IKK、JNK、ElF2α、インスリン受容体などのいくつかの炎症性キナーゼと相互作用することにより、この応答に寄与します。この代謝的に活性化された炎症性複合体は、代謝インフラマソームまたはメタフラマソームと呼ばれます。PKRは、ミトコンドリアの小胞体ストレス応答(UPR mt)にも関与しています。ミトコンドリアの小胞体ストレス応答では、PKRは転写因子AP-1を介して誘導され、PKRを活性化できるPACTとは独立して活性化されます。ヘルペスウイルスはPKRが炎症により殺されないための様々な機構を身に付けました。ヘルペスなどは、デコイdsRNA、分解、ウイルスdsRNAの隠蔽、二量体化ブロック、基質の脱リン酸化、または偽基質によってPKRの働きから逃れることができるのです。 たとえば、エプスタインバーウイルス(EBV)は、自分の遺伝子EBER1を使用しておとりdsRNAを生成します。これが、バーキットリンパ腫、ホジキン病、鼻咽頭癌、およびさまざまな白血病などの癌を作り出してしまうのです。おとりdsRNAとは何でしょうか? プロテインキナーゼR(PKR)は、自然免疫経路の重要な抗ウイルス成分であり、EBウイルスなどのウイルスの二本鎖RNA(dsRNA)によって活性化されます。他にアデノウイルスが作り出すRNA1は、PKRに結合することによっておとりとして機能しますが、PKRの活性化は誘導せずに、逆におとりRNA1(VAI)によってPKRの抗ウイルス応答を阻害します。RNA1は多く存在する非コードウイルスRNAです。おとりRNA1(VAI)では、高親和性PKR結合を仲介する拡張ヘリックスが生成されますが、PKRを短すぎてアクティブ化できません。アデノウイルスと同様に、エプスタインバーウイルスは高濃度の非コードRNA、EBER1を生成します。PKRとおとりRNA1(VAI)およびEBER1との相互作用を比較しますと、両方のRNAは、dsRNAを介したPKR活性化の阻害剤として機能します。ただし、EBER1はPKRを弱くアクティブにしますが、VAIはアクティブにしません。PKRはVAIよりも弱くEBER1に結合します。生理学的イオン濃度でのアッセイは、両方のRNAが2つのPKRモノマーに対応し、PKR二量体化を誘導できます。
PKRに対するウイルス防御メカニズム
| 防御タイプ | ウイルス | 分子 |
| デコイ(おとり)dsRNA | アデノウイルス | VAI RNA |
| エプスタインバーウイルス | EBER | |
| HIV | TAR | |
| PKRの劣化 | ポリオウイルス | 2A pro |
| ウイルスのdsRNAを隠す | ワクシニアウイルス | E3L |
| レオウイルス | σ3 | |
| インフルエンザウイルス | NS1 | |
| 二量体化ブロック | インフルエンザウイルス | p58 IPK |
| C型肝炎ウイルス | NS5A | |
| 疑似基質 | ワクシニアウイルス | K3L |
| HIV | Tat | |
| 基質の脱リン酸化 | 単純ヘルペスウイルス | ICP34.5 |
リン酸化PKRおよびeIF2α(Eukaryotic Initiation Factor 2、略してeIF2)がアルツハイマー病(Alzheimer’s disease、略してAD)患者の海馬および前頭皮質の変性ニューロンで活性化しています。それはアルツハイマーの原因は海馬がヘルペスウイルスに感染して傷つき崩壊した時に、細胞内から放出されたDAMP moleculesを処理するためにPKRが活性化されるためです。さらに、これらのニューロンの細胞周辺に見られる、リン酸化タウタンパク質に対するIgM抗体が見られるのはDAMP molecules(細胞傷害関連分子パターンの分子)に対してBリンパ球が一時的に作った抗体がIgM抗体であります。活性化されたPKRは、細胞質および核に特異的に見られ、ニューロンのアポトーシスマーカーと共に局在していたのはDAMPの処理をするのがPKRであるからなのです。血中および脳脊髄液中にPKRよりもリン酸化PKRが多いのはPKRを活性化してDAMPの処理をする必要があるからです。 ADの患者では、異常な高さの総PKRおよびpPKRレベルが見られるのは神経細胞である海馬の細胞に感染したヘルペスが脳細胞を崩壊させた後のDAMPによって生まれた分子(molecules)を処理するために、PKR-eIF2αアポトーシス促進経路が活性化してヘルペスウイルスのみならずヘルペス感染細胞を殺したり、傷つけたりした結果、海馬の神経細胞が変性し、神経病理学的病変を引き起こすにも関わらずPKRがアルツハイマーの責任の一端を担っているなどというアホなことをいう医学者が多すぎます。原因と結果をさかさまにしているからです。いくら結果を原因だと思い込んで研究しても全て無駄です。
PKRの活性化は、アルツハイマー病患者のBACE1(β部位APP切断酵素)発現の抑制解除を介してアミロイドβペプチド(Aβ)の蓄積を引き起こされるのはヘルペスが海馬の細胞に感染したために正常な働きが出来なくてBACE1が発現できなくなるからです。APPとは、英語でamyloid precursor proteinで略してAPPで、訳してアミロイド前駆体タンパク質です。BACE1とは、β-secretase(β- セクレターゼ)とも言い、BACE1 の英語はbeta-site APP cleaving enzyme 1です。BACE1は、ニューロンでアミロイドβペプチドを生成するための主要なベータセクレターゼ(酵素)の役割を果たしています。つまり酵素であるBACE1(β-secretase)が活性化するとアミロイドペプチドが産生されてアルツハイマーが生ずると言われていますが、これは大間違いです。アミロイドβはアルツハイマーの原因ではないからです。通常、BACE1プロモーターの5’非翻訳領域は英語で5’untranslated region、略して5’UTRは、BACE1(β-secretase)遺伝子の発現を基本的に阻害します。なぜかというと、ヘルペスが感染している神経細胞がDAMPの moleculesになることは絶対にないからです。ところが実際にヘルペス感染が起こってしまうとBACE1の発現が必要となってしまうのです。BACE1の発現によってeIF2a(Eukaryotic Initiation Factor 2a)がリン酸化されて活性化され、BACE1の5’UTRによって発揮される阻害効果が逆転し、阻害効果がなくなってアルツハイマーが生ずると言われていますが、これは大間違いです。なぜならばeIF2aのリン酸化はヘルペスウイルスが感染した海馬の細胞の処理のためにはPKRの活性化が必要であり、PKRが活性化するとBACE1の阻害効果がなくなります。まさに単純ヘルペスウイルス(HSV)や酸化ストレスなどを起こすヘルペスウイルスによる細胞感染は、PKR-eIF2a経路の活性化を通じてBACE1の発現を増加させ、傷ついた細胞をPKRによって処理するのです。さらに、BACE1の活性の増加はAPP-βCTFの合成につながります。APP-βCTFは英語でC-terminal fragment of APP, βCTF、別名C99 AD(Alzheimer’s disease)のエンドソームのβ-アミロイド前駆体タンパク質の非常に活性の高い(APP-βCTF)誘発性機能障害を起こすβ部位を切断されたカルボキシ末端フラグメントを生み出すのです。エンドソームは、非常に活性の高いβ-アミロイド前駆体タンパク質β(APP-βときにβ-APPとも書く)のプロセシング部位(処理部位)であり、β-アミロイド前駆体タンパク質のβ部位を切断してしまいC-terminal fragment of APPを作ってしまうのです。ヘルペス感染細胞の傷害により異常になったエンドソームは、初期のエンドソーム抑制因子であるrab5というタンパク質の発現のアップレギュレーション(増加)を起こしてしまいます。以上は、AD(Alzheimer’s disease)における最も初期の既知のアルツハイマーに特異的な海馬のニューロン応答です。まずBACE1の活性の増加は、APP-βCTFの合成につながり、次にβCTFのレベルが上昇すると、rab5の過剰活性化が引き起こされ、βCTF(C-terminal fragment of APP、別名C99)は、APPL1は英語でAdaptor Protein, Phosphotyrosine Interacting With PH Domain And Leucine Zipper 1といい、アダプタータンパクとして様々なタンパク質を集め、様々な役割があり、APPL1はrab5があるエンドソームに動員され、そこでAPPL1はrab5をGTPに結合させて活性化します。GTPは英語でguanosine triphosphateで、訳してグアノシン三リン酸で、略してGTPで、細胞内シグナル伝達やタンパク質の機能の調節に用いられ、Rab5タンパク質は、GTPと結合することによって活性化され、GDP結合によって非活性化される状態を交互に繰り返すのです。GTP-rab5は病理学的に加速されたエンドサイトーシス、エンドソームの腫れ、およびrab5エンドソームは神経の軸索輸送を選択的に障害され、神経細胞の働きに異常が起こるのです。これも元はと言えば、神経細胞に感染したヘルペスウイルスが原因なのです。
GTPとGDPとは何でしょうか? GTPは、英語でguanosine tri-phosphateで、訳してグアノシン三リン酸、略してGTPは、ヌクレオチド(核酸)であり、正式名はグアノシン-5′-三リン酸です。グアノシン二リン酸 (guanosine di-phosphate、略してGDP) からアデノシン三リン酸 (ATP) のリン酸を受容して生合成されます。類似した構造を持つ ATP が生物体内で高エネルギーリン酸結合のエネルギーを利用して、様々な生合成や輸送、運動などの反応に用いられるのに対し、GTP は主として細胞内シグナル伝達やタンパク質と結合してタンパク質の機能の調節に用いられます。GTPは、GTPアーゼの作用によってグアノシン二リン酸(GDP)に変換されるセカンドメッセンジャーメカニズムにおいて、特にGタンパク質を用いたシグナル伝達に不可欠です。
GTPが関わる生合成反応ではRNA合成やその他ヌクレオチドの合成に用いられ、多糖の生合成では中間産物であるGDP-グルコース、GDP-マンノースなどグアノシン二リン酸-糖とも言われるGDP糖の合成に用いられます。言うまでもなく、GDPはグアノシンヌクレオシドのピロリン酸エステルです。つまり、GDPはピロリン酸基、五炭糖のリボースそして核酸塩基のグアニンから構成されています。
GTPと結合するタンパク質を下に掲載しておきます。これらは各機能の1サイクルごとに GTP 1分子と結合し、GDP に加水分解したのち解離します。
1)タンパク質翻訳で働くいくつかの因子: 翻訳開始因子、翻訳伸長因子(アミノアシル tRNA のリボソームへの結合、ペプチジル tRNA の転座)、翻訳終結因子(真核生物)。
rRNAとは何でしょうか? 今をときめくmRNAのワクチンはSARS-Cov-2のSタンパクmRNAを鋳型としてスパイクタンパク質をまず作らせることですね。このようにmRNAを鋳型としてタンパクを作る段階を翻訳(translation)といいます。ところがタンパク質の生合成にはmRNA以外に、トランスファーRNA(tRNA)やリボソーム(ribosome)が必要です。原核細胞と真核細胞の翻訳機構は大変良く似ているのは、生命にとって重要な機構は生物種を超えて保存されてきたからなのです。タンパク質合成の場であるリボソームは大小2つの粒子よりなり、真核細胞では4種のrRNAと82種のタンパク質から成り立ち、rRNAは触媒活性をも持っています。これをribozyme活性といいます。rRNAは英語でRibosomal ribonucleic acid、訳してリボゾームリボ核酸です。真核細胞では、リボソームは小胞体膜に結合し、粗面小胞体を形成します。一方、ミトコンドリアやペル-オキシソームや核で必要なタンパク質は遊離の状態のリボソーム(free ribosome)でつくられます。
リボソームリボ核酸(rRNA)は、すべての細胞に不可欠なリボソームの主成分である非コードRNAの一種です。 rRNAは、リボソームでタンパク質合成を行うリボザイム(リボ酵素)です。リボソームRNAはリボソームDNA(rDNA)から転写され、リボソームタンパク質に結合して大小のリボソームサブユニットを形成します。 rRNAは、リボソームの物理的および機械的因子であり、力ずくでトランスファーRNA(tRNA)とメッセンジャーRNA(mRNA)を協力させてmRNAの情報をタンパク質に翻訳させるのです。リボソームRNAは、細胞に見られる様々な全てのRNAの中で最も多い主要なRNAで細胞のRNAの約80%を占めています。もちろんrRNA自身はタンパク質に翻訳されることはありません。 リボソームは、質量で約60%のrRNAと40%のリボソームタンパク質から成り立っています。
翻訳伸長因子のアミノアシル tRNA のリボソームへの結合とは何でしょうか?アミノアシルtRNAとは何でしょうか? アミノアシルtRNAは英語でamino-acyl-tRNAです。アミノアシルtRNAはtRNAのアデノシン残基中の五単糖の3′のヒドロキシ基(OH)とアミノ酸のカルボキシル基(COOH)がエステル結合して生じる分子で、生体内では各アミノ酸に対し、1 つのアミノアシル tRNA 合成酵素が存在します。アミノアシルtRNA合成酵素 は英語でamino-acyl-tRNA synthetaseで、略してaaRSやARSで、特定のアミノ酸をそのアミノ酸に対応するtRNAにエステル結合させてアミノアシルtRNAを合成する酵素です。アシル基とは、カルボン酸からOHを抜いた形、すなわちR-CO-というような形の基であり、エステル結合というのは、カルボン酸とアルコールから成るカルボン酸エステル (carboxylate ester)であり、カルボン酸エステルの特性基 (R−COO−R’) をエステル結合 (ester bond) と呼びます。ちなみに、エステル結合による重合体はポリエステル (polyester) と呼ばれ、多価カルボン酸(ジカルボン酸)とポリアルコール(ジオール)とを、脱水縮合してエステル結合を形成させることによって合成され、石油から作られる素材の一種で、衣料品にはポリエステル繊維として使われます。ポリエステル繊維には大きく分けて3種類あり、一般的なのはPETと呼ばれるもので、ペットボトルの原料としても有名です。PETで作られるポリエステル繊維は、軽くて耐久性が高いのが大きな特徴です。
さて本論に戻りましょう。リボソームに運ばれてきたtRNA部分の3塩基からなるアンチコドンが、mRNAのコーディング領域のコドンと対合し、タンパク質合成が始まります。従って、3塩基のコドンと1アミノ酸の対応づけが行われる場はリボソームであっても、実際にコドンとアミノ酸の対応関係を示す遺伝暗号はaaRS(amino-acyl-tRNA synthetase)の特異性に基づいています。通常の生物では翻訳に使用されるアミノ酸20種類に対し、それぞれ対応するaaRSをもっています。例えば、アルギニンを認識してアルギニンtRNAにエステル結合する反応を触媒するaaRSはアルギニルtRNA合成酵素 (arginyl-tRNA synthetase) のように表記され、略号はArgRSのようにアミノ酸3文字表記+RSです。
翻訳伸長因子のペプチジル tRNA の転座 とは何でしょうか?まずペプチジル(基)とは何でしょう? ペプチジルはペプチドの形容詞です。したがって「ペプチドの」という意味ですが実際はペプチドはアミノ酸から出来ていますから、ペプチドの末端の「アミノ酸の」という意味と考えておいて下さい。真上の文章でアルギニンというアミノ酸がアルギニルという言葉に変わっているのは「アルギニンの」という意味です。さてここでかつてリボソームはmRNAに書き込まれた遺伝情報を忠実にタンパク質に翻訳するための装置であるとイメージされていましたが、実はリボソームは細胞内の状態に応答して機敏に遺伝子発現を制御する情報処理装置であるのです。リボソームは細胞質の状態を検知しながら、いわば細胞質にどっぷりと浸かって遺伝子情報をタンパク質への翻訳を行っています。
リボソームにおける翻訳伸長過程は大きく分けて,1)コドンの解読,2)新たなペプチド結合を形成するペプチド転移反応,3)次のコドンへのリボソームの転座の3つから成り立っています。ペプチド転移反応の活性中心であるペプチジルトランスフェラーゼが存在するセンター(PTC)は大サブユニットにあり,新たに合成されたペプチドは,大サブユニットを貫く出口トンネルを通って出てきます。出口トンネルはおよそ100Åの長さがあり,“伸びた”状態の新生ペプチドでは30~40アミノ酸残基を保持しています。出口トンネルの内壁は大部分がrRNAで形成されていますが,出口トンネルに突き出た狭窄部位があります。

下にペプチド転移反応の活性中心(PTC)と新生ペプチドの通り道である出口トンネルと狭窄部位を示します。ペプチド転移反応によってP部位のペプチジル–tRNAからA部位のアミノアシル–tRNAへとペプチドが転移され,ペプチドが外れたP部位のtRNAがE部位に移動し,アミノ酸残基がひとつ伸びたA部位のペプチジル–tRNAがP部位へとそれぞれ移動します。なお,A, P, Eの各部位を示すために3つのtRNAを描いていますが実はtRNAが3つ同時に存在することはないのです。
2)チューブリン: GTP 結合型が重合して微小管を形成する。GDP に加水分解すると脱重合する。
3)7TM受容体からのシグナル伝達に関わるGタンパク質: GTP を結合した状態が活性型で下流にシグナルを伝達する。
4)低分子 GTP 結合タンパク質:Rasタンパク質など、シグナル伝達や細胞機能の調節に働く。Gタンパク質と同様に GTP を結合した状態が活性型として働く。
5)グアニル酸シクラーゼ: これもシグナル伝達に関与するタンパク質である。GTP からサイクリックGMP(cGMP) を合成し、cGMP はセカンドメッセンジャーの一種として機能する。グアニル酸シクラーゼには心房性ナトリウム利尿ペプチド受容体(膜結合型)や、一酸化窒素によって活性化される可溶型のグアニル酸シクラーゼが知られています。
リン酸化PKRは、ヘルペス感染が生じている病的なニューロン(神経細胞)にリン酸化タウタンパク質と一緒に見られます。アルツハイマーには必ずリン酸化タウタンパク質もアミロイドβ(Aβ)と一緒に出現します。元来タウ-タンパク質(Tau protein)は中枢神経系および末梢神経系の神経細胞(ニューロン)やグリア細胞に発現しているタンパク質で、微小管結合タンパク質(Microtubule-associated protein、略してMAP)のタンパク質の一種であり、微小管の重合や安定化を維持しています。また微小管以外にもさまざまなタンパク質と結合しており、生後の脳の成熟、軸策輸送およびそのシグナル伝達の調節、熱ストレスに対する細胞応答、成体での神経発生など、脳神経系で起こるさまざまな現象を調節しています。ところがこのタウ-タンパク質がリン酸化することがアルツハイマー病などの神経変性疾患の原因となると考えられました。そこで脱リン酸化を触媒するプロテインホスファターゼ1と2Aの酵素を阻害するオカダ酸(OA)という薬剤を投与して正常なタウタンパクのリン酸化を阻害するとどのような結果になるかを調べました。すると、このオカダ酸(OA)はタウタンパクのリン酸化、Aβ(アミロイドβ)沈着および神経細胞死を増加させました。しかも、さらにこのオカダ酸(OA)はPKRリン酸化の誘導するどころか、eIF2aリン酸化も誘導してしまいました。すると次に、このeIF2aのリン酸化により、活性化された転写因子4(activating transcription factor 4、略してATF4)が誘導され、アポトーシスと転写因子4(ATF4)の核移行が誘導され、神経細胞死をもたらしてしまいました。さて、以上の研究は何を意味するのでしょうか?タウタンパクをリン酸化させないことによってアルツハイマーの実験的なモデルを作ったつもりなのでしょうが、実はもっと素晴らしい結論が隠されていたのですが研究者はその真実に全く気づいていないのです。OAは単なるプロテインホスファターゼ1と2Aの酵素を阻害するオカダ酸(OA)は同時に他の生命に必要なプロテインホスファターゼの働きも阻害してしまっていたのです。その結果、細胞は薬剤性DAMP moleculesを生み出してPRRであるPKRを活性化しリン酸化させてしまったのです。
PKRがリン酸化するのはDAMPによって生じた分子を処理するためです。AD脳(Alzheimer’s disease 脳)では、活性化されたPKRとGSK3βの両方がニューロン(神経細胞)のリン酸化タウと共に存在しているのは、グリコーゲンシンターゼキナーゼAβ(GSK-3β)は正常なタウをリン酸化させ、さらにアポトーシスを含むいくつかの細胞機能に関わっています。
Ⅰ型インターフェロンのIFN-α、IFN-βを活性化するのにPKRが関わっています。 Ⅰ型インターフェロン(IFN-α、IFN-β)応答は,免疫応答の数時間前から数日前に働く防御機構であり,ヘルペスウイルスの侵入,転写,翻訳の開始などを妨げます。インターフェロン応答としてはdsRNA依存的タンパク質キナーゼ(PKR)が関与する経路と2′-5′-オリゴアデニル酸合成酵素(2’,5′ oligoadenylate synthetase、略して2-5AS)が関与する経路の主に2つの経路があります。これらの経路の活性化は細胞内すべての遺伝子発現の低下を引き起こし、細胞の機能を低下させ、アポトーシスに導くのです。
なぜ私のような若年性アルツハイマーが起こるのでしょうか?老人性アルツハイマーと若年性アルツハイマーの違いは何でしょうか?それはBACE1 による APP 切断部位選択機構の違いによって生じるのです。
アルツハイマー病(AD)は、アミロイドβタンパク質(Aβ)が可溶性オリゴマーを形成しシナプス毒性を発現することで発症すると言われていますが、海馬の細胞に感染したヘルペスウイルスを感染細胞もろとも殺し、隣接する細胞に感染しないように人体の免疫が働いているだけでシナプス毒性を発現することによってAβ(アミロイドβ)が生じるのではないのです。Aβは、元来正常な人のI 型膜タンパク質であるアミロイドβ前駆体タンパク質(APP)が細胞外領域でアスパラギン酸プロテアーゼで I 型膜タンパク質であるβセクレターゼ(BACE1)による切断を受け、さらに細胞膜内でγセクレターゼ複合体による切断を受けることでAβ(アミロイドβ)が産生されます。BACE1 (β-secretase)による APP の切断部位として Asp-1 (アスパラギン酸1の部位)と Glu-11(グルタミン酸11の部位) の 2 か所であります。番号は Aβ(アミロイドβ)配列のアミノ酸部位を示します。Asp-1 切断からは Aβ1-40 を、Glu-11 切断からは Aβ11-40 が産生される。AD 患者脳内では、何らかの要因によ り Asp1 の切断が増加し、Aβ1-40、及び凝集性の高い Aβ1-42 が増加することが AD 発症の原 因の一つであると考えられている。若年性 AD 患者において切断部位である Glu11 の変異が 発見され、この変異体では Glu11 の切断が抑制され、Asp1 における切断の増加、それに伴い Aβ1-40 および Aβ1-42 の増加が認められる。
