やっとやっと終わりが見えてきました。ここで最後なので最後まで着いて来て下さい!!!
初めての方はPart1から読んで下さい!
さて統合失調症の原因となっているドパミンについて詳しく見ましょう。又何故、ドパミンが、過剰になり何故、過剰なドパミンが統合失調症を引き起こすのかの答えを少しずつ出していきましょう。例のごとく難問ですよ!!!
まずドパミンを産生・分泌するドパミン作動性神経はどのようにドパミンを合成するのでしようか?
ドパミンは、必須アミノ酸ではないチロシンからチロシン水酸化酵素によってL-ドパに、L-ドパはドパ脱炭酸酵素によってドパミンに合成される。更にドパミンはドパミンβ水酸化酵素の働きで、ノルアドレナリンに合成される。ノルアドレナリンはCOMT(カテコール-O-メチル転移酵素)によってノルメタネフリンへ、最後にMAO(モノアミン酸化酵素)によって、MHPG(3-メトキシ-4水酸化フェニールグリコール)へと分解され代謝されてしまいます。
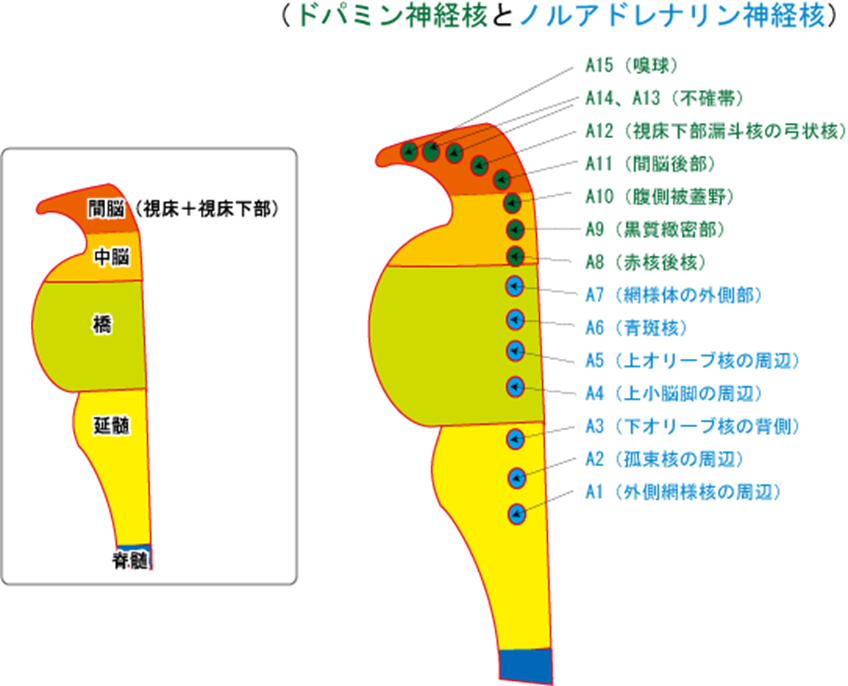
統合失調症の原因を作るドパミン作動性の神経核について詳しくみましょう。下図に脳幹の間脳と中脳と橋と延髄にあるドパミン神経核とノルアドレナリン神経核の存在部位を正確に示す絵を掲載します。統合失調症は天才でもなりうるので知性の病気ではないのです。つまり大脳新皮質の病気ではないのです。感情や情動が統合失調症という病気にかかわっているのです。感情と情動のちがいはなんでしょうか?感情は情動を含み、情動は動物的な感情です。因みに研究者によっては間脳を脳幹に含めない人もいます。従って脳幹は中脳と橋と延髄の三つの部位から成り立っているという人もいます。
大脳辺縁系は食欲や性欲などの生存本能や恐怖や好き嫌いなどの人間の本能的な感情や情動を司る旧皮質と古皮質の二つの皮質を含みます。旧皮質は大脳皮質のうち、系統発生的に最も古い部分をいう。古皮質とは海馬、歯状回、脳弓から成り立ち海馬、歯状回は記憶に関り、脳弓は大脳辺縁系の複数の領域をつないでいます。哺乳類では、旧皮質は古皮質とともに大脳辺縁系を形成し、食欲・性欲などの本能行動や、情動行動を司っています。古皮質は大脳皮質の一部で、系統発生的には旧皮質に次いで現れる部分です。両生類以上に見られ、両生類では旧皮質とともに古皮質は脳の主要部を占める。哺乳類は魚類から、両生類へ、そして爬虫類を経て最期に誕生しました。
脳の進化は「大型化」の歴史です。その働きを減らすことなく再編成しながら利口に大きくなっていくために、作り直すのではなく新たなる部位を付け加えるリフォームつまり増改築しながら益々賢く大きくなって最後に人間の脳にまで進化したのです。因みにセキツイ動物は「哺乳類・鳥類・爬虫類・両生類・魚類」の5種類に分類され、無セキツイ動物は 「節足動物(昆虫類/甲殻類/クモ類/多足類)・軟体動物・その他」と分類されます。
ヒトの大脳皮質では、新皮質が発達し、皮質の90%を占めることになりました。新皮質が外側をおおってしまったため、残りの皮質である古皮質と原皮質は、大脳半球の下とか奥のほうに追いやられてしまいました。旧皮質と古皮質からできている大脳辺縁系は他の動物も持っている本能的な情動に関り、人間にとっても最も基本的な食欲や性欲、快か不快のような本能的な情動を司るのが大脳辺縁系の役割です。
大脳の新皮質が大きくなったため、鳥類以下の動物での運動中枢だった神経細胞の塊(かたまり)も、大脳の底に追いやられました。これが、大脳基底核ですね。錐体路という、哺乳類に特有の運動中枢が出現したため、残った基底核は、下位中枢になりました。残った大脳基底核は、なめらかかつしなやかな運動ができるように、錐体路の中枢を補佐しています。
大脳辺縁系は、脳梁に沿って、かつ大脳の奥深くにある線条体と言われる尾状核、被殻からなる大脳基底核を取り巻くように存在する古い皮質の総称です。主な役割は、記憶(短期記憶と長期記憶)と情動(やる気、怒り、喜び、悲しみなどの快不快)に関することです。大脳辺縁系を構成(こうせい)するものは、眼窩回、透明中隔、帯状回、海馬傍回、内嗅皮質、海馬体、扁桃体、乳頭体です。特に帯状回、海馬傍回、海馬体、扁桃体は重要です。その中でも偏桃体は「情動の中枢」ですから好き嫌いや快不快という最も原始的かつ動物的な情動がこの偏桃体から生まれるのです。戦争の原因は偏桃体の存在なのです。快楽を得るために弱肉強食は民主主義と言われる資本主義国は戦争をし続けるでしょう。悲しいですね。
因みに眼窩回とは(Orbital gyri)は、前頭葉の脳回。同義語に前頭連合野眼窩部、眼窩前頭前野、前頭前野眼窩部です。回とは哺乳類の大脳半球の表面には『しわ』すなわち凹凸(おうとつ)が存在するが、その『しわ』の隆起部分を脳回といい、脳回の横にある陥凹(かんおう)部分を脳溝という。脳には多くの脳回と脳溝が見られる。
海馬傍回(Parahippocampal gyrus)は海馬回(hippocampal gyrus)とも言い海馬の周囲に存在する灰白質の大脳皮質領域です。大脳内側面の脳回のひとつである。この領域は記憶の符号化及び記憶の検索において重要な役割を担っている。海馬傍回はあくまでも大脳皮質であって結して海馬の一部ではないのです。
乳頭体は視床下部にある左右1対の半球状の高まりで,その内部に嗅覚に関係する乳頭体核がある。
嗅内皮質は海馬と共に記憶に深く関わる脳領域であり、双子のように類似した構造を持つ2つの領域、内側嗅内皮質と外側嗅内皮質から構成されます。 内側嗅内皮質は空間情報を処理する一方、外側嗅内皮質の細胞は匂いや物体の情報処理に関わります。臭いは、その人の過去の経験とつながっています。納豆のような強い臭いの食べ物でも、美味しいと感じる体験を一度すれば、「美味しい臭い」という記憶になるのです。臭いが記憶と結びつきやすいのは、脳の中で臭いと記憶を担当する場所「嗅覚野」と「海馬」)がとても近くにあるからです。
人間はほかの動物と比べ新皮質の占める割合が著しく高くなっていますが進化的に古い大脳辺縁系の割合は他の動物と比べるとさほど変わらないのです。従って本能だけについて言えば人間は動物並みと言えます。アッハッハッハ!しかし偏桃体から快・不快の情報を受け取った帯状回ではその情報を行動に結び付けるかの判断を下すのですがその行動を実際に実行するかどうかの最後の決定は帯状回から行動意欲の情報を受け取った新皮質が決めるのです。
大脳辺縁系の偏桃体は人間の本能である「情動の中枢」であり好きか嫌いか、快か不快かという原始的な動物的な感情が極めて自然に生まれるのが偏桃体です。何故好きか嫌いか、快か不快かという理由は偏桃体にとってはどうでもいいのです。しかしその偏桃体の情動(感情)に基づいて快不快を実行に移すかを決めるのは大脳の新皮質であるのです。しかし大脳皮質も間違いをしでかすことが多いのは人間の大脳皮質も大したことがないと言えるでしょう。あっはっは!!!
何故人間は38億年にわたる長い長い時間をかけて進化した最高の大脳新皮質をもっているのにもかかわらずいつまでも同じ多すぎる間違いを犯すのでしょうか?それは真実を知るために人間は進化したのではなく自己の快楽を最大限に獲得するために利害・損得で大脳皮質も最終決定を下すからです。しかも一度、強い快楽を味わって大脳皮質に長期記憶として残されてしまうとつよい欲求が生まれてしまうのです。ついでに言えば最後に増改築され加えられた「脳の中の脳」である前頭連合野をサルと比べると、サルでは大脳皮質の12%を占めるのに人では約30%ですからこれが人と猿の大きな違いを生み出したのでしょうが心はサルと変わらないどころかサル以下ですね。悲しいですね。
快楽を求める人間は一番最初に生れて初めて快楽を記憶として残し始める経験は何だと思いますか?又生まれてすぐに必要なものは何だと思いますか?空腹感と喉の渇きをいやすことです。栄養としての食べ物と水分です。つまり母乳です。此処から人間は快楽の経験とその快楽の記憶と欲望の第一歩を踏み始めるのです。これこそ意識と記憶の芽生えであり母乳が美味しいと記憶されるのでお腹が減りだすと泣いて母乳を要求する欲求を母親に求め始めるのです。泣きわめくと欲求が満たされ快楽と言う報酬が得られるという記憶がますます強い記憶として残ります。これは原初的な学習と言えます。しかも空腹を癒すというこの快楽経験の中での母親とのやり取りから言葉の記憶が開始され言葉が記憶され人間のみに許された言葉のやり取りが可能になるスタート点に立つことが出来るのです。ことばの習得も快楽の充足から生まれるのです。
これを医学的に説明すると空腹と言う内的な情動の刺激や生まれてすぐの血糖の低下や塩分の低下などの情報は情動を支配する大脳辺縁系に取り込まれ腹がすいたからミルクが飲みたいというという衝動を生み出します。この衝動は空腹から生まれた欲求とミルクを飲んで空腹が満たされた欲求の充足により幸せを感じることが出来るのは快楽の中枢である側坐核からドパミンが出され100%満足して幸せを感じ安らかに眠りの付くことが出来るのです。
脳幹のドパミンの神経群として重要なのが、A8:赤核後核とA9:黒質緻密部とA10:腹側被蓋野の三つです。特にA9:黒質緻密部とA10:腹側被蓋野の二つは統合失調症と極めて関りがあるので重要です。もちろんA11~A15までのドパミンの神経群も少ないながらドパミンを産生しているので統合失調症の長い経過の中で症状に影響が及ぼされるのは当然です。
| 神経集団番号 | 神経集団名 | 主な作用 |
| A8 | 赤核後核 | 辺縁系へ投射 |
| A9 | 黒質緻密部 | 新線条体へ投射。パーキンソン病の関連部位として錐体外路運動に関与 |
| A10 | 腹側被蓋野 | 辺縁系へ投射する中脳辺縁系路と前頭葉へ投射する中脳皮質路がある。前者は扁桃体の興奮(情動)により活性化され、後者はストレスや不安等負の要因で活性化。 |
| A11 | 間脳後部 | 視床下部へ投射。レストレスレッグス症候群の関連部位 |
| A12 | 弓状核 | 下垂体ホルモンの分泌を制御する。この系の抑制はプロラクチンの分泌を促進する(D2ブロッカーの高プロラクチン血症の原因部位)。 |
| A13、A14 | 不確帯 | 視床下部へ投射 |
| A15 | 嗅球 |
上の表のドパミンの主な作用について解説しましょう。
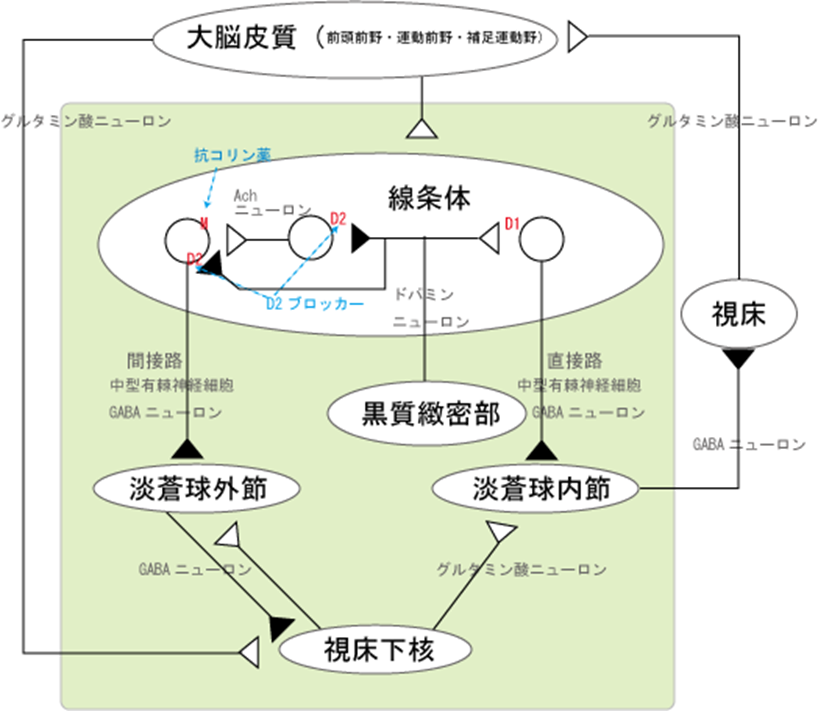
A9:黒質-線条体路におけるドパミンの作用は黒質-線条体路では、中脳の黒質(A9)に細胞体を置くドパミンニューロンが、線条体(被蓋、尾状核)でGABAを神経伝達物質とする抑制性の中型有棘神経細胞を抑制的に制御しています。ドパミンは運動機能、意欲および快感に関連する行動を担っている神経伝達物質です。脳が興奮すると線条体(被蓋、尾状核)と呼ばれる特定の脳領域でドパミンが大量に放出されます。線条体にはドパミンD1受容体(D1R)を発現する中型有棘神経細胞(D1R-細胞)とドパミンD2受容体(D2R)を発現する中型有棘神経細胞(D2R-細胞)の異なる2種類の神経細胞が存在します。D1Rは、プロテインキナーゼA(PKA)と呼ばれるリン酸化酵素を活性化し、逆にD2RはプロテインキナーゼA(PKA)の働きを抑制します。PKAは細胞の興奮性や報酬(快感)関連行動に関係していることから、ドパミンはPKAを介してD1R-細胞の興奮性を高め、D2R-細胞の興奮性を抑制すると考えられてきました。この時少なくなったドパミンはPKAを介してD2R-細胞の興奮性を抑制制御することが出来なくなった上にドパミンの働きを抑えるアセチルコリンが働いたらどうなるでしょうか?
この抑制制御がうまく行かない上に、線条体へのドパミン量が低下する事によりアセチルコリン神経の活性化によりGABA神経の興奮 → 視床下核の興奮 → 視床の抑制で錐体外路障害を生じるのがパーキンソン病。脳内の神経伝達物質の一つアセチルコリンはドパミンと互いに拮抗し合う物質である。一般的に、脳内のドパミン作用が弱くなっている状態ではアセチルコリンの作用が強くなっている。アセチルコリンの働きを抑える作用を抗コリン作用といって、この作用によってドパミンの作用を強めることができる。アセチルコリン神経の活性化によりGABA神経の興奮→視床下核の興奮→視床の抑制→錐体外路障害???を生じるのがパーキンソン病となります。視床下核とは、大脳基底核を構成する核のひとつであり、大脳皮質から興奮性入力を、淡蒼球外節から抑制性入力を受け、淡蒼球外節・内節、黒質網様部に興奮性投射を送る。視床下核が障害を受けるとヘミバリスムを来す。
近年、パーキンソン病に対する脳深部刺激療法(DBS)のターゲットとして臨床的にも注目されている脳内のアセチルコリンは大脳皮質や海馬などに広く分布して神経を興奮させ記憶力や学習意欲などを高める。脳内のアセチルコリンは、目覚めさせる(覚醒)作用や活力を上げる(賦活)作用を持ち、重要な神経伝達物質(神経細胞の突起先端部のシナプス部で放出されて、次の神経細胞に情報を届ける重要な役目を持つ物質)です。このアセチルコリンを作る細胞がアルツハイマー型認知症の脳で減っています。作る方の神経細胞は増やせませんので、シナプスでのアセチルコリン分解消失を抑えることでアセチルコリン減少を補う薬剤が、アルツハイマー型認知症治療薬として我が国で最初に開発されました。それがドネペジル(商品名アリセプト)です。その後、同様な働きを持つガランタミン(商品名レミニール)やリバスチグミン(商品名イクセロンやリバスタッチ、貼り薬)が開発され、現在はこの3剤が日本で使われています。レビー小体型認知症でもアセチルコリンの減少があり、アリセプトはレビー小体型認知症でも認可されています。これらの薬は、どちらかというと「元気系」です。ごく一部の方ですが、活発になりすぎたり、頻繁に怒るようになったり、介護者が困る症状が出てくることがあります。これは効き過ぎ症状ともいえるので、効き過ぎの場合は薬を減量~中止することで、介護者が困る症状は改善します。
この3剤の副作用は、第一に胃腸障害です。アセチルコリンによって胃液(胃酸)がたくさん分泌され、嘔気・嘔吐・食欲低下などを引き起こします。また、腸の動きが良くなりますので、下痢も副作用です(便秘が良くなるという恩恵もありますが)。
視床下核は、グルタミン酸作動性の投射ニューロンにより構成されている。長い樹状突起を有し、また軸索は核内走行中に軸索側枝を出す。大脳皮質からグルタミン酸作動性の興奮性入力を、淡蒼球外節からGABA作動性の抑制性入力を受け、淡蒼球外節・内節、黒質網様部にグルタミン酸作動性の興奮性投射を送る三つの経路があります。直接路・間接路モデル・ハイパー直接路(hyperdirect pathway)です。
1. 直接路(direct pathway):線条体から直接、淡蒼球内節・黒質網様部に到る経路
2. 間接路(indirect pathway):線条体から、淡蒼球外節、視床下核を順に経由して淡蒼球内節・黒質網様部に到る経路・ハイパー直接路(hyperdirect pathway)
直接路・間接路モデルは、それまで入り組んでいた大脳基底核の神経回路を、整理し明快にまとめたばかりでなく、後から述べるように大脳基底核疾患の病態や定位脳手術の治療メカニズムも説明できる画期的なものであった。これによって、視床下核は間接路の重要な中継核と位置づけられた。 さらに、視床下核も大脳皮質から直接、入力を受けていることが以前よりわかっていたが、最近では大脳皮質からの入力が特に重要という認識が広がり、線条体と並んで大脳基底核の入力部と考えられるようになった。それに伴い、直接路・間接路に加えられたのが、三つ目のハイパー直接路(hyperdirect pathway)です。
3. ハイパー直接路(hyperdirect pathway):大脳皮質から入力を受けた視床下核ニューロンが淡蒼球内節・黒質網様部に投射する経路
視床のイメージができにくいので視床ゲート機構といわれる末梢からの嗅覚以外の他のすべての感覚情報が視床を経由する際に、上位中枢である大脳皮質へ送る情報を選択する機構についてみましょう。視床ゲート機構においては抑制性細胞の集合体である視床網様核が中心的な役割を担っています。感覚系視床において主に取り上げられることが多い概念だが、運動系視床も基本的に同じ構造の回路を持ち、同様のゲート機構であります。
視床ゲート機構とは嗅覚を除く感覚系の情報は視床を通るときには視覚の情報は視床の外側膝状体を通り、聴覚の情報は、視床の内側膝状体を経由して側頭葉の聴覚野へ送る。因みに幻視、幻聴、妄想、認知機能障害(統合失調症)の患者には、視床のドパミントランスポーター(DAT)の機能に変化があり、健常者より30%のDAT増加がみられるとの報告があります。一方、視床のドパミンD2受容体の量が減少しているとの報告もあり、視床のドパミン神経系の過剰活動が、情報の統合に乱れを生じさせていると考えられる。また、振戦の原因部位と考えらえられています。パーキンソン病薬(L-dopa、ドパミンアゴニスト)の過剰な服薬により幻視、妄想が出現するとの報告もあります。
パーキンソン病の運動症状の出現は、Braak仮説によると、抗α-シヌクレイン抗体を用いて高齢者の中枢神経系におけるLewy小体の分布を詳細に検討、Lewy小体はまず嗅球に出現、迷走神経背側核(延髄)、視床と、その後、下部脳幹(橋)、中脳黒質、扁桃体へ上行進展して発現します。扁桃体に優位にLewy小体が分布しているとの報告もある。
視覚の情報は視床の外側膝状体を経由して大脳皮質の視覚情報を受け取り後頭葉の一次視覚野へと送られる。一次感覚野とは大脳皮質において感覚情報が入力される最初の部位で,視覚,聴覚,体性感覚といったそれぞれの感覚について一次感覚野が存在し,そこで感覚情報の内容に応じた情報の選別がなされ,異なる特徴をもつ情報は異なる脳領域においてさらなる情報処理が進む.われわれはそういった一次感覚野のはたらきのおかげで入力された感覚の特徴を認識し,快感あるいは不快感を得たり,感覚入力に対し咄嗟の行動を起こしたりすることができる。すべての情報がそのまま伝達されるわけではなく、必ず視床において情報が選別されているのは視床ゲート機構に果たす視床網様核の役割は、視床網様核からの抑制性入力によって、大脳皮質へ送る情報と送らない情報を選別する機構があるからです。聴覚の情報は視床の内側膝状体から聴放線を通り側頭葉の聴覚野に送られる。
因みに外側膝状体(lateral geniculate nucleus略してLGN、又は,lateral geniculate body略してLGB)は、脳の視床領域の一部であり、中枢神経系の網膜から情報を受け取り、視覚情報の処理を行う。 LGNは網膜から視神経、視交叉、視索を通って直接情報を受け取る。一方、内側膝状体(Medial geniculate body略MG)とは視床に属する神経核群であり、中脳下丘と大脳皮質聴覚野の間に位置する聴覚伝導路の中継核である。上行系は下丘から、下降系は大脳皮質聴覚野から入力を受ける。大脳皮質聴覚野へ送る聴覚情報の選別が内側膝状体の主な機能である。
視床で大脳皮質へ送られる情報(必要な情報)と送られない情報(不要な情報)が選別されている。選別には視床網様核からの抑制性入力が中心的な役割を担っている。嗅覚を除く感覚系の情報は視床(例:外側膝状体(視覚)、内側膝状体(聴覚))を経由して大脳皮質一次感覚野へと送られる。この際、すべての情報がそのまま伝達されるわけではなく、視床において情報が選別されている。視床網様核からの抑制性入力によって、大脳皮質へ送る情報と送らない情報を選別する機構があると考えられている。
脳弓は大脳辺縁系の複数の神経核の領域をつなぐ仕事をしています。
D2受容体には、D2L受容体とD2S受容体のサブタイプが存在している。
D2L受容体は、辺縁系では後シナプス、線条体では中型有棘細胞の後シナプスに多く発現している。D2S受容体は自己受容体として黒質、腹側被蓋野のドパミン神経終末(前シナプス)に発現している。
一方で、中脳皮質路のドパミン神経においては自己受容体が欠如しているため、ストレス刺激によって活性化してしまうとドパミンの分泌が制御できなくなるという。
統合失調症では、主として前頭葉のNMDA受容体やドパミン受容体の異常によってドパミンの分泌が亢進し、大脳基底核の黒質や線条体のD2受容体を刺激してしまうため、視床が興奮して、ちょっとのことでもビクッとしてしまうような知覚過敏の状態を作り出してしまいます。統合失調症の原因はドパミンが過剰に作られというよりもドパミン受容体の異常によってドパミンの分泌が亢進し、大脳基底核のD2受容体を刺激してしまうため言われるが嘘です。基本的には長期のストレスのためにA8A9A10のドパミン神経核がドパミンの分泌を亢進させ過ぎたからです。
統合失調症のドパミン仮説とは何か?
統合失調症の病態に関する仮説の一つで、精神疾患の病態仮説としては最も長く精力的に検証が行われてきた仮説の一つである。このドパミン仮説の最も確実な根拠は、統合失調症の症状を軽減するのに有効な抗精神病薬の共通の特徴がドパミンD2受容体のアンタゴニストである点である。 初めに提唱されたドパミン仮説は、統合失調症の病態はドパミン神経機能の過活動、とするものであった。私が今証明しようとしているのは 統合失調症の病態は統合失調症の患者さんが脳幹に存在する二万足らずのドパミン作動性神経機能の過活動、とりわけ脳が分泌するドパミンの90%近くを占めるA8,A9,A10のドパミン作動性神経の過活動をストレスによって15年以上もかけて起こし続けたためにドパミン過剰になったのが統合失調症の原因の一つなのです。その後、統合失調症学会は前頭葉のドパミン神経機能の低活動性を伴う大脳皮質下のドパミン神経機能の過活動、と修正してしまいました。この文言も矛盾が合い過ぎません?ストレスホルモンが過剰になると高等な脳の前頭前野が委縮することが分かったからです。結局この修正も間違いでした。さらにドパミン以外の多くの病因が引き起こす共通病態としての新たなるドパミン仮説が捉え直されるようになり今日に至っていますが今も尚、「統合失調症のドパミン仮説」は統合失調症についての真実は何も語られてはいません。
真実は 「患者さんがストレスに対して脆弱でありすぎたので患者さん自身が長年かけてストレスがかかるたびにステロイドホルモンと同時に過剰にドパミンを出しすぎたからです。かつherpesがとりわけ脳の情動や快楽を支配する大脳辺縁系の扁桃体に増えたためです。」
現代の統合失調症の(不完全な一部の真実しか語らない)ドパミン仮説はどのような経過で生まれたのでしょうか?統合失調症の病態に関する仮説の一つがドパミン仮説であり、精神疾患の病態仮説としては最も長く精力的に検証が行われてきた仮説の一つですが、 この仮説の最も確実な根拠は、統合失調症の症状を軽減する現在用いられている有効な抗精神病薬の共通の特徴がドパミンD2受容体のアンタゴニスト(拮抗剤)である点である。薬物受容体と相互作用して細胞内シグナル伝達を引き起こす物質をアゴニスト(作動剤),アゴニストと拮抗的に作用してその作用を減弱させる物質をアンタゴニスト(拮抗剤)と呼ぶのは既に述べました。
最初に提唱されたドパミン仮説は、統合失調症の病態はドパミン神経機能の過活動、とするものであったが、その後、前頭葉のドパミン神経機能の低活動性を伴う皮質下のドパミン神経機能の過活動と修正されたこともすでに述べました。。
皮質下とは大脳のどこを指すのでしょうか?「大脳皮質下領域の神経細胞体が集簇している核」を略して皮質下と言います。大脳の深部にある構造で、大脳辺縁系の海馬、扁桃体の二つ、大脳基底核の側坐核、尾状核、被殻、淡蒼球の三つ、視床、視床下部などが含まれ、系統発生的に古い領域です。運動機能や記憶・情動・意欲などに関与します。このように下等な動物も大脳皮質が十分発達していなくても、優れた運動機能も必要な記憶力も感情も強い生きる意欲も持っている証拠である系統発生的に古い領域は発達して保持しているのです。この様に下等な動物でも言葉は無いのに記憶があるのは大脳皮質が進化していなくても五感の感覚神経で覚えるからです。
次に出てきたドパミン仮説は多くの病因が引き起こす共通病態としてドパミンが絡んでるという仮説が捉え直されるようになり今日に至るまで二転三転したのにもかかわらず今なお統合失調症の真実の原因は不明のままです。その答えを私が見つけた統合失調症の真実の原因は三つあります。すでに述べたように一つは快楽を求めすぎる生まれつきの気質、つまり快楽のホルモンであるドパミンを欲しがる気質、二つ目はストレスがかかりやすい気質、三つめはその結果、ヘルペスが脳内に増えたからです。とりわけ脳の 大脳辺縁系の海馬、扁桃体、大脳基底核の側坐核、尾状核、被殻、淡蒼球と視床、視床下部などの系統発生的に古い領域にherpesが増殖してしまったからです。
確かに上に挙げた三つの原因のために大脳皮質下のドパミン神経機能の過活動は正しいのですがその過活動の原因についてはこのドパミン仮説は一言も言及していないのが弱点です。もう一度、統合失調症の真実の原因を繰り返すと「この統合失調症になり易い生まれつきのストレスに弱い優しい気質を持って生まれ、かつストレスの多い環境で育って免疫を下げた。」のが一番目の原因です。二番目の原因はやはり「ストレスのたびごとにherpesが増え知らぬ間に私と同じように徐々に脳に感染しだした。」からです。三つ目の原因は「ストレスを感じるたびごとに副腎皮質でステロイドホルモンを、かつ副腎髄質で快楽ホルモン(神経伝達物質)であるドパミンを必ず作り出してしまった。」からです。この三つの要因である生まれつきの気質と増えたherpesと増えたドパミンに焦点を合わせて統合失調症の原因を説明すればすべてのタイプの統合失調症は原因をはじめ症状と治療が説明可能となり統合失調症は実は治せる病気であると理解できるのです。さースタートしますよ!!!
まずドパミンは脳のどこで、かつどの神経核で、かつどの神経細胞で産生され、かつどの神経細胞からどの部位にある神経にドパミンは投射されるのかを説明しましょう。
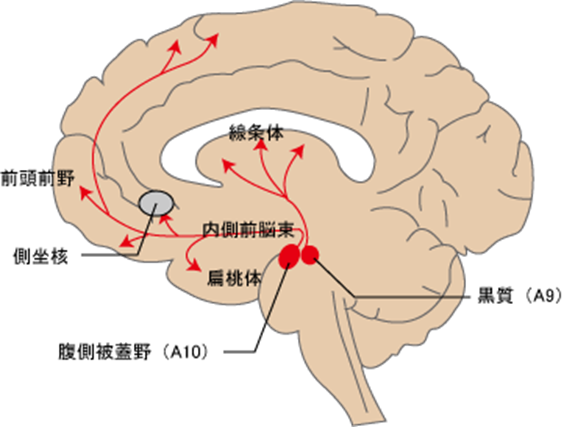
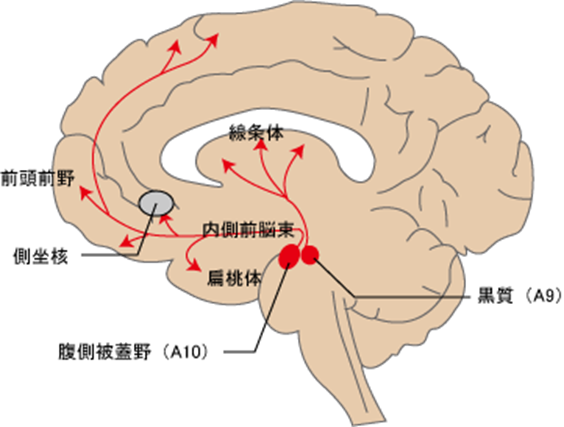
まずA10ドパミン作動性神経核の腹側被蓋野の二つの投射系路の中脳辺縁系路と中脳皮質路とは何か?から話を始めましょう。 A10ドパミン作動性神経核は「快感中枢」と言われます。
中脳腹側被蓋野から投射されるドパミンニューロンは大きく、腹側被蓋野-辺縁系路(側坐核・海馬・扁桃体等)と腹側被蓋野-皮質路(前頭前野)の2つに分けることができます。
一つ目の腹側被蓋野-辺縁系路は快情動、報酬系に関わる情動回路を動かすドパミンを分泌していやな思い(ストレス)をのりこえれば快楽を報酬として手に入れられる経路であり、長い間自分の思いを遂げられない落胆ばかりを繰り返すと快楽は手に入れることが少ないので益々、ストレスが溜まるばかりで徐々に統合失調症の症状が出始めます。いわゆる陽性症状の発現に関わることになります。
卑近な例を挙げますと腰痛の痛みで悩んでいる人が誰かに腰をさすってもらったら痛みが収まった経験を快い情報として「情動の中枢」である扁桃体が感じ認識すると扁桃体自身も感覚の記憶として覚えて置き、かつ価値ある記憶としては「記憶の中枢」である海馬が大事に忘れないように収納してくれます。この快さや快さの感覚と記憶をまず味わうのは扁桃体です。快楽ホルモンと言われるドパミン作動性の神経集団であり 「快感中枢」であるA10を活性化すると大量のドパミンが分泌されこのドパミンが 「幸福の神経核」の側坐核を興奮させて、視床内背側核から大脳皮質の 「快楽の終着点の前頭前野」に伝わり快楽を味わうことのできる Yakovlevの情動回路を再び作動させて、もういちどさすって貰いたい!さすってもらえば痛みが収まる!といった快の情動行動を起こす方向へと進むのですが、さすってくれる人がいなければ快さが得られないとまたこれがストレスになります。このように 「快感中枢」であるA10神経核は情動を司る脳幹や大脳辺縁系を支配しています。
A10神経核は中脳辺縁系路にA10神経核自身が分泌したドパミンを自分自身が受容する多数のドパミンレセプターを持っておりこれを自己受容体とかオートレセプターと言います。
ドパミンの自己受容体は何のために生れたのでしょうか?
D2受容体には、D2L受容体とD2S受容体のサブタイプが存在している。
D2L受容体は、辺縁系では後シナプス、線条体では中型有棘細胞の後シナプスに多く発現している。D2S受容体は自己受容体として黒質、腹側被蓋野のドパミン神経終末(前シナプス)に発現している。
一方で、中脳皮質路のドパミン神経においては自己受容体が欠如しているため、ストレス刺激によって活性化してしまうとドパミンの分泌が制御できなくなるので中脳皮質路のドパミン神経においては自己受容体が欠如しているため、厳しい現実と欲望にまみれた思いの現実とを正確に区別できなくなり自分の世界に閉じこもることになってしまうのがSZのヒトなのです。
自己受容体とかオートレセプターは何の目的で生まれたのでしょうか?ストレス刺激によってドパミンは活性化してしまうのでドパミンの分泌が制御できなくなると長年にわたってストレスがかかりやすい気質の人は快楽に関わる思いと現実との見分けがつかなくなって統合失調症のような病気にならないためにブレーキをかけているのです。しかしすべてのドパミン神経に快楽を抑制してくれる 自己受容体がありすぎてドパミンが減ってしまうと快楽は少なくなり、ドパミン神経に快楽を抑制してくれる 自己受容体が多すぎると、快楽の欲望が生まれつき強すぎるすべての人はSZになってしまうでしょう。ワッハッハ!!!人間と言うのは本当に厄介な存在ですね。だからこそ中庸が重んじられるのです。
セロトニン神経にもオートレセプターという機構が備わっています。セロトニン神経が興奮すると、オートレセプターを介して自己にセロトニン神経の働きを抑制するネガティブフィードバックをかけ、セロトニン神経の増えた活動を抑えてしまうのです。このため、簡単にはセロトニン神経の活動レベルは高く維持し続けられないようになっているのです。セロトニン神経の オートレセプターをセロトニンで 絶えず繰り返し刺激し続けてやると、オートレセプターに結合するセロトニンが結合する オートレセプターの数が減少していきセロトニンの作用も正常になってしまうのです。
腹式呼吸法などのリズム運動によって、セロトニン神経が活性化するのには、3か月かかる。セロトニン神経の自己受容体の数が減少して、自己の抑制を解放するためであるらしい。
うつ病やパニック障害や不安障害、心身症の人が、仏教カウンセリングで腹式呼吸法や坐禅をこころみる場合にも、指導を受けた後、毎日、実践してもらう必要がある。最低2カ月もすれば、症状が緩和されたのを実感できる。心身症とは、身体疾患の内、発症や経過に心理社会的ストレスの影響で機能的(器質的)な障害を伴った疾患群です。うつ病の人が、腹式呼吸法、自己洞察法(坐禅に似た心理療法)を開始して快楽を求める生活を中心を控えて自分の心を常に見て快楽の感情に動かされない生活を心がけることです。このように自分で自分にカウンセリングし始めると、突然、「気分の好転」や「穏やかな並み幸せ」を感じることが出来ます。
自分中心の生活から一歩も二歩も引いて自分の心を省察し反省していると、ドパミン神経のみならずセロトニン神経の オートレセプターの数が減ることで、それで、最終的にはドパミン神経の活動のみならずセロトニン神経の活動レベルが高く維持されるという。こうなると、力を入れなくても、坐禅も楽になるし、日常いらだち事、感情の爆発が少なくなるだろう。もう少し続けて6カ月~1年ほどたつと、心の病気は治癒し、セロトニン神経が根本的に活性化しているから、再発の可能性が低くなる。ストレスに反応しすぎることも少なくなり、他の心の病気、心身症やがんの発病の確率も低くなるだろう(ストレスは免疫にも影響する)。
セロトニン神経には「自己受容体」があり、自己が自己を抑制する。そして、その抑制を解放するのは、知性によらずかつ快楽によらず、できる限り自分を離れて生きれば自分のストレスホルモンは必要なくなるのです。人の永遠の心のしんの幸せは言い換えると自分の心の貪欲に惑わされずできる限り自分を忘れる心と自分を忘れる生活にあることに気付くことです。
ドパミンが通る経路の途中にはA9神経(黒質)とおなじ視床下部の他に側坐核や大脳辺縁系に属するあらゆる神経核や側頭葉の灰白質の神経細胞があり、 最後の終着点はあらゆる精神的な知的快楽をはじめとする能力的な金銭的な快楽を味わうことができる前頭連合野の数多くの神経細胞が終着点となります。脳幹の神経の神経線維は無髄神経線維です。というのは有髄神経線維であるとヘルペスが髄鞘にも感染してしまうと髄鞘から神経線維に感染し神経線維から移動して神経細胞体まで感染してしまうからです。 生命中枢が多い脳幹の神経核はたった2万個しかなくしかもその神経線維が無髄であるのでヘルペスが感染しにくいのです。生命中枢が多い脳幹の神経核にherpesが感染してしまうと生命を常に危険にさらすことになるのでヘルペスが脳幹の神経細胞に感染しにくいようになっているのです。言い換えると神経細胞に感染するのは末梢神経の髄鞘のシュワン細胞や中枢神経の髄鞘を形成する神経膠細胞から軸索(神経線維)の感染して更に軸索を順行性や逆行性に長い時間をかけて移動して中枢神経の神経細胞の神経細胞体にまで感染していくのです。という間接的な証拠になるのです。
セロトニンが「幸せ」の脳ホルモンと言われるように「幸福の神経核」と言われる側坐核はドパミンの受容体を大量に持ち快感・報酬を求める行動や意欲の向上に重要な役割を果たし向上心やる気や行動力をもたらし、かつ「脳の中の脳」である前頭連合野を援助して向学心や知的勉学行動意欲などをもたらす一方、最高の快楽と幸福を求めすぎるのでアルコールや覚せい剤の嗜癖ももたらすのです。
さてその側坐核はどこにあるのでしょうか?両側の大脳半球に一つずつ存在し、大脳基底核と大脳辺縁系の境界に位置し、尾状核と被殻が、透明中隔の外側で接する場所にあるので大脳基底核と大脳辺縁系のいずれにも属していると考えられているのですが一応、大脳基底核に位置しています。従って、 側坐核は嗅結節などとともに腹側線条体の一部であるのです。実はこのように大脳基底核と大脳辺縁系は曖昧なのでこのような言葉と曖昧な概念はなくした方が混乱を招かないという学者がたくさんいます。私も賛成です。
モルヒネなどは、体内に取り込まれると腹側被蓋野でドパミン神経のレセプターと結合して刺激し、側坐核へ投射する快感中枢神経である10神経細胞群の末端からドパミンの分泌を促し、投射する神経とのシナプス間隙のドパミンが増えることにより、シナプス後細胞が興奮状態となって、モルヒネ摂取者は「何ものにも代えがたい幸福感」を味わい、モルヒネに対する精神的な依存が形成され、モルヒネを止めることは極めて困難となります。
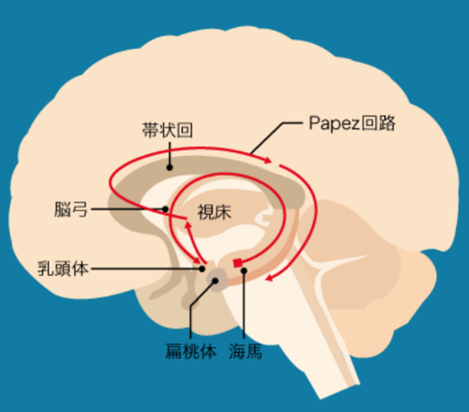
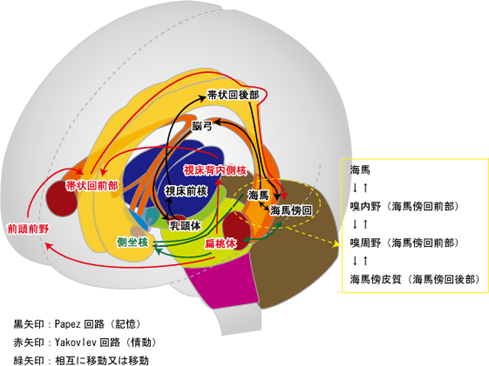
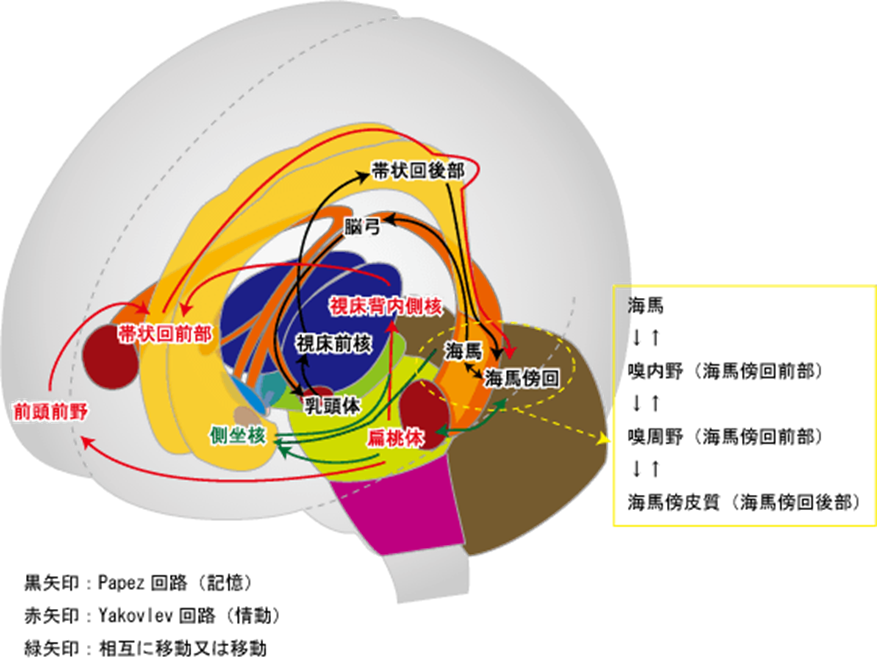
上図に主に記憶の神経回路であるパペッツの回路と主に情動のヤコブレフの神経回路の二つの回路を同時に示していますが、この図の中の緑矢印の説明は「相互移動又は移動」と書かれていますが意味が不明確ですから説明します。下図をよく見てください。側坐核から出ていく緑の経路と偏桃体から出ていく赤い経路とが融合していますが互いに独立して神経活動をしていることを意味しているのです。
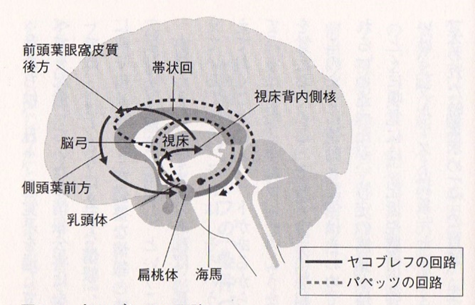
「幸福の神経核」の側坐核に注目してもらいたいのです。パペッツの回路とヤコブレフの回路の二つの回路の両者とも側坐核に神経回路がつながっていることは情動(快楽)にも市の快楽の記憶にもかかわっていることを意味しています。
二つ目の腹側被蓋野-皮質路のほうは、投合失調症になってしまうと不安やストレスによりA10ドパミン作動性神経核が活性化し、負の行動を起こす陰性症状の発現に関わっています。
何故、辺縁系の側坐核に行くドパミンでは陽性症状が出て大脳皮質の神経にドパミンが向かってしまうと陰性症状が出てしまうのでしょうか?それは大脳皮質が進化したのは本能である快楽を味わうためではなく快楽を統御するためですからドパミンに対する反応は下等な辺縁系の細胞が過剰なドパミンに対して反応する仕方が高等な大脳皮質の細胞とは異なるのは当然なのです。両者の神経細胞の遺伝的な反応の違いが症状の違いとして出るのです。
繰り返しますが統合失調症には、健康なときにはなかった状態が表れる陽性症状と、健康なときにあったものが失われる陰性症状があり、 陽性症状の典型は、幻覚と妄想です。 幻覚の中でも、周りの人には聞こえない声が聞こえる幻聴が多くみられます。 陰性症状は、意欲の低下、感情表現が少なくなるなどがあります。
腹側被蓋野から皮質にいく経路路へは中脳から大脳に投射するドパミン神経が中枢のドパミン神経系の大部分を占め、大脳皮質は運動機能、認知機能などの中枢機能の調節に関与します。また、ドパミン神経系は統合失調症の精神疾患の病態生理に対する関与しているので、抗精神病薬等の治療薬や依存性薬物の標的となる。小胞内に貯蔵されたドパミンは開口放出によって細胞外に放出される。ドパミン神経の投射部位のみならず黒質や腹側被蓋野でもドパミンは放出される。これらの部位では細胞体や樹状突起からドパミンが放出され、特に黒質ではそれが主であると考えられている。
軸索終末からの放出も細胞体・樹状突起からの放出も共にCa2+依存性であるが、軸索における放出の方がより高濃度の細胞外Ca2+を必要とする。線条体においてドパミン放出部位と考えられる構造の60-70%は明確なシナプス構造を形成していない。また、ドパミン受容体の大部分はシナプス外の部位に発現している。従ってドパミンによって担われる信号伝達は、主として放出部位から比較的離れた受容体に作用する拡散性伝達(Volume transmission)によるものである。
ストレス刺激が起こるとステロイドホルモンとドパミンとノルアドレナリンが分泌される上に、ドパミンがA10経路を活性化してしまい、さらに多くのドパミンが分泌されるので、前頭前野のドパミンD1受容体を介して前頭前野の行う認知・思考・ワーキングメモリ等の機能を抑制してしまいます。快楽が強すぎると前頭前野の行う認知・思考・ワーキングメモリ等の理性的な機能が発揮できないのは当然です。
Papez回路(記憶)記憶は脳内のネットワークによって担われています。記憶の重要な回路として知られているのがPapezパペツ回路です。Papez回路は大脳辺縁系の海馬を中心とした神経ネットワークです。海馬から脳弓を経て、乳頭体、視床、帯状回を通って海馬にもどります。脳弓は大脳辺縁系の複数の領域をつなぐ。健忘を起こす脳の部位のほとんどがPapez回路にふくまれています。帯状回は大脳辺縁系の各部位を結びつける役割を果たしており、感情の形成と処理、学習と記憶と情動(感情)に関連した回路としてヤコブレフ回路が知られている。
Yakovlevヤコブレフ回路という扁桃体を中心とした回路もあります。この回路は、情動や感情に関係する神経ネットワークとして知られています。
嬉しい、楽しい、悲しいのような情動に関する記憶は、むかしのことでもよく憶えています。海馬と扁桃体は隣どうしにある部位で、おたがいに密接に関係し、Yakovlev回路とPapez回路が相互作用することで、感情的な記憶がよく記憶されるのではないかと考えられています。記憶に関わる神経回路はPapez回路と呼ばれています。帯状回が興奮する事で海馬 → 海馬采・脳弓 → 乳頭体(乳頭体視床路を通る)→ 視床前核 → 帯状回 → 海馬傍回 → 海馬体を結ぶ経路で持続的に興奮する事で情動が生まれ記憶に関与する事が知られている。(パペッツ回路)情動・本能などに関与するほか、恐怖・攻撃・性行動・快楽反応にも関与。記憶に関連する他の脳部位として、前脳基底部のマイネルト基底核、ブローカ対角帯、内側中隔核かあり、アセチルコリン神経(ACh)の起始核(acetylcholine産生部位)です。
Yakovlev回路(情動)Yakovlev回路:側頭葉皮質前部(38野) → 扁桃体 → 視床背内側核 → 前頭眼窩皮質 → 鉤状束 → 側頭葉皮質前部―偏桃体という回路を形成。鉤状束(こうじょうそく、英: Uncinate fasciculus)は、大脳にある連合線維で、外側溝の下部を横切り、 前頭葉の脳回と側頭葉の前端を繋いでいる。Yakovlevの回路は、扁桃体、視床背内側核、帯状回前方、前頭葉眼窩皮質後方、側頭葉前方、偏桃体から成る障害の基本像は,行動異常と健忘症状です。健忘については,Papez(ペーペズ,パペッツ)回路とともに,いずれがどのように関与しているかの詳細はまだよく分かっていないようです。Yakovlev(ヤコブレフ)回路とは,大脳辺縁系で情動と記憶に関与する中枢です。
統合失調症で問題になるのは知能ではありません。異常な感情や情動による苦しみです。それではまず正常な感情や情動がどのようにして生まれるのでしょうか?人間として当たり前の感情や情動は大脳辺縁系の偏桃体で生み出される脳内ホルモンと呼ばれるモノアミン系で分泌される神経伝達物質によって大脳に運ばれます。心の感情の情報を例えば喜怒哀楽などの感情の情報を偏桃体で判断して大脳に伝える神経伝達物質はホルモンに似ていますので脳内ホルモンと名付けられたのです。この脳内ホルモンの分泌を仕切っているのはモノアミン系と言われる神経細胞の集団です。このモノアミン系は脳幹に沿って存在するたった2万個の神経細胞の集団なのです。モノアミン系は役割によってA,B,Cの3つの系列に分かれています。A列はノルアドレナリンを分泌するA1~A7とドパミンを分泌A8~A12の神経細胞に更に区分されます。A6のノルアドレナリンを分泌する神経細胞の集団を「怒りの中枢」と言います。A10のドパミンを分泌する大きな細胞集団は人間にしかないのです。一方、C列からはアドレナリンが分泌されB列はA列のホルモンの分泌を抑えかつC列のホルモンの分泌を抑制する役割を担っています。これらの脳内ホルモンが脳内に伝わって緊張や興奮や快楽などの生理的感情的変化を身体にもたらすのです。感情は脳内ホルモンの分泌によって引き起こされるのです。
セロトニンはノルアドレナリンやドパミンの暴走を抑えて、心の落ち着きや安定感をもたらす神経伝達物質である。セロトニンの分泌が減少すると、人はやる気や集中力が低下し、イライラしてキレやすくなったり、寝つきが悪く不眠になったり、気持ちが落ち込んでうつ状態になったりする。しかし、うつ病の治療のためにセロトニン作動系の抗うつ剤を多量に服用したり、薬剤の相互作用によって脳内のセロトニン濃度が高まり過ぎると、今度は、発汗や心拍数の増加、吐き気、筋肉の痙攣、体の震え、頭痛、錯乱、昏睡などの、いわゆる「セロトニン症候群」と呼ばれる症状が出現する。
つまり、脳内ホルモンは、増えすぎても減りすぎても心身のバランスが崩れて、人は容易に不安定な状態に陥ってしまうのである。
セロトニンは必須アミノ酸トリプトファンから生合成される脳内の神経伝達物質のひとつです。 視床下部や大脳基底核・延髄の縫線核などに高濃度に分布しています。 他の神経伝達物質であるドパミン(喜び、快楽など)やノルアドレナリン(恐怖、驚きなど)などの情報をコントロールし、精神を安定させる働きがあります。
ドパミンの自己受容体
D2受容体には、D2L受容体とD2S受容体のサブタイプが存在している。
D2L受容体は、辺縁系では後シナプス、線条体では中型有棘細胞の後シナプスに多く発現している。 D2S受容体は自己受容体として黒質、腹側被蓋野のドパミン神経終末(前シナプス)に発現している。
一方で、 中脳皮質路のドパミン神経においては自己受容体が欠如しているため、ストレス刺激によって活性化してしまうとドパミンの分泌が制御できなくなるという。
快楽の王者と言ってよい側坐核の特徴を勉強しましょう。
側坐核は大脳線条体の前腹側部に位置する神経核であり,その神経の発達的特徴や解剖学的特徴は,線条体と共通点が多く,線条体システムの一部 と考えられる。一方で,特に側坐核のシェルとよばれる部位の神経細胞は、扇桃体 より入力を受け,視床下部に直接軸索をのばすなど、いわゆる拡張扇桃体の発生的および解剖学的特徴も共有する。これらのことから,側坐核には拡張扇桃体と背側線条体との移行部分としての機能が見られる。側坐核には扇桃体,海馬体の一部である海馬台をはじめとした辺縁系の線維以外に前頭前野,前頭眼窩などの皮質性線維,視床の背内側核、さらには脳幹の ドパミン,セロトニン、ノルアドレナリン作動性線維などが収赦する。また,側坐核は上述 したシェル部 とコアと呼ばれる部位よりなる。シェルとコアは入出力様式が異なる。
例えばラットの下辺縁(infralimbic)皮質は主にシェルに,前辺縁 (prelimbic)皮質の背側部は主にコアに投射し,海馬台からの線維は主にシェルに投射する。側坐核は側坐核-線条体と腹側被蓋野-黒質赦密部を結ぶ大規模なスパイラルネットワークの起始部でもある。腹側被蓋野と黒質繊審部よりなる中脳ドパミン系は側坐核や線条体と双方向性の神経結合を形成している。このことは側坐核の活動が拡散し最終的には運動野の活動を制御する可能性を示している。さらに,彼らはこの大規模な双方向性の回路について同様に側坐核と深い関わりを持つ大規模なスパイラルネットワークが存在する。側坐核シェルは,前頭前野の腹内側部 より入力を受け取り,腹部淡蒼球を介して,視床内側核背部の内側に投射する。視床内側核背部の内側部の神経は前頭前野腹内側部に線維背外側部に出力し,この部位からは側坐核コアに投射がある。コアから派生する線維は腹部淡蒼球の別の部位に到達し,視床背内側に至る。さらに,これから派生する線椎も,閉回路を形成しつつ拡散 し運動野に至る。
側坐核の役割は何でしょうか?解剖学的な側坐核の特徴は,いわゆる情動系と運動系の接点としての重要性を示している。一方で,その機能に関しては主に快楽報酬系としての役割がある。例えば,側坐核は腹側被蓋野のドパミン神経より強い投射を受けるが,エンケファリンやコカインなどのシナプス間隙内のドパミン量を増大させる薬物を自己投与すると自己投与行動の頻度が増大する。また,自然報酬獲得のための接近行動が側坐核のドパミン拮抗物質の投与やドパミンの枯渇によって減弱するのは,側坐核ドパミンシステムが報酬獲得の役割があることがわかる。
視床前核群: 前端の視床前結節を形成し、三つの核群(前背側核、前腹側核および前内側核)からなる。乳頭体から線維をうけ、帯状回の皮質と結合する。大脳辺縁系と関連しており、情動や新しい記憶と関連すると考えられている。
視床内側核群[背内側核]: 内側髄板の内側を占める核群である。大脳半球の葉、視床下部、大脳核の線条体などと連絡し、体性、内臓性情報を統合して前頭葉へ投射する。感覚に基づく情動に関係し、この部分が侵されると強い不安状態に陥る。また、視床下部から自律神経系にも作用する。
前腹側核:淡蒼球、前頭葉運動領と連絡する。
外側腹側核: 上小脳脚からの線維を受け入れるとともに、前頭葉の運動領とも連絡する。
後腹側核: 体性感覚の中継核で後内側腹側核および後外側腹側核がある。後内側腹側核は三叉神経毛帯と孤束核からの味覚線維を受入れ、後外側腹側核は内側毛帯や脊髄視床路の線維を受入れ、中心後回に連絡する。末梢からの体性知覚の中間中枢をなす重要な核である。
視床外側核群: 内側および外側髄板の間にある核群で、体性知覚の中間中枢をなすとともに錐体外路系に属する。視床外側核群は背側外側核と腹側核に分けられる。背側外側核は辺縁系と関連し、記憶や情動形成に関わる。後外側核は視床の他の核からの線維を受け、頭頂連合野に連絡する(上頭頂小葉)。これにより、感覚情報は連合野で解析・統合される。
視床後核群: 視床後端部の視床枕と内側および外側膝状体を形成する核群である。視床枕は皮膚知覚、視覚および聴覚の統合に関与する。内側膝状体は外側毛帯と上丘からの線維を受入れ、聴放線を側頭葉の聴覚中枢に送る。外側膝状体は視索からの線維を受け入れ、視放線を後頭葉の視覚中枢に送る。
正中核群: 網様体あら線維を受け、大脳皮質を覚醒して意識を明晰に保つ上行性網様体賦活系に関わる。室傍核、中心核複合核および結合核からの遠心性線維は扁桃体複合核に投射する。視床の正中核群の中のいくつかは、帯状回の皮質の前部にも投射しているかもしれない。また、有髄、無髄の線維が視床下部と連絡していると考えられる。
遺伝子で決まるどんな気質の人が、また気質遺伝子の発現と環境によって決まるどんな性格の人が統合失調症には多いのか、またなり易いのか?
また、 控えめ・内気・おとなしい人や神経質な部分と無頓着な部分を持ち合わせている人、人からの言動等に傷つきやすいといった性格の人、人とコミュニケーションが苦手な人が統合失調症の患者さんには多いといわれています。
統合失調症にかかる原因として生まれながらの様々な素因とストレスの大きさが関係していると考えられます。
ストレスの大きさというのはストレス自身の大きさとそれを受け止める人のストレスに対する耐性で定義してみるとわかりやすいかもしれません。
すなわち同じストレスがあってもストレスに敏感すぎる人はそれによって心のバランスを崩した結果、さまざまな不調を起こしやすく、ストレスに対して上手に対処する方法を身に着けている人は心のバランスを崩しにくく心も体も大きな影響を受けることが少ないと考えられます。
統合失調症の発症原因の1つに過剰なストレスがあるので、ストレスの対処法を身につけることも大切です。
なりやすい要因、タイプ
ストレスを受けやすいタイプ
統合失調症になりやすい要因として、遺伝的要因、脳のトラブル、気質・性格などの要因があり、一方で強いストレスにさらされると発症するというのが、先にあげたストレス・脆弱性モデルです。
この仮説に立てば、ストレスを受けやすい人は、そうでない人よりも発症の可能性は高いということになります。長崎大学が行なった調査では、統合失調症発症者のうち、約80%の人が3ヵ月以内に入学、就職、失恋、結婚、死別など環境の変化があったことがわかっています。
もともと脆弱性を抱えている人でストレスに弱い人が、長期にわたってストレスのかかる出来事を体験すると、統合失調症を発症する確率は高くなるものと考えられています。
なりやすい性格
統合失調症と性格に関しては、クレペリン(ドイツの精神科医)、ブロイラー(スイスの精神科医)、クレッチュマー(ドイツの精神科医)などが、性格に一つの傾向が見られることを指摘しています。ただし、これはあくまで傾向です。自分の性格と照らし合わせて不安がる必要はないのですが、参考までに列記すると以下のようになります。
・男性では無口、内気、ひきこもりがち、孤独
・女性では怒りっぽく、敏感、神経質、強情
・易怒性と引っ込み思案、孤独な性格。
・内向的で非社交的、内気、孤独で生真面目
・臆病、恥ずかしがり、敏感、神経質
・従順、正直、鈍感
近親者に統合失調症の人がいる
統合失調症と遺伝の関係は、無視できません。
統合失調症の患者を対象とした調査では、以下のような結果が報告されています。
・親の片方が統合失調症であった場合、子どもが発症する確率は10%。
・両親がともに統合失調症であった場合、子どもが発症する確率は40%。
・統合失調症の兄弟姉妹がいた場合、当人が発症する確率は約10%。
確かに近親者に統合失調症の人がいれば、そうでないケースよりも発症の確率は高い。しかし、一方で、こんな調査報告もあることを忘れないようにしましょう。
・統合失調症の患者さんの家族のうち、両親の約9割は統合失調症ではない。
・統合失調の患者さんのうち、兄弟を含めてもその約8割は統合失調症ではない。
・統合失調症の患者さんのうち、甥や姪を含めても約6割は統合失調症ではない。
なりにくい要因
ストレスをうまく発散できる
ストレスは、統合失調症に限らず様々なタイプの精神疾患の呼び水となっています。ストレスに弱い人は、精神疾患にやられる可能性が高くなります。
といって、それは生まれつきの性格ともからんでいますから、ストレスに強くなろう、といっても無理なところがあります。
しかし、ストレスをうまく処理するテクニックは習得できるはずです。自分の性格にフィットした方法で、ストレスを発散する手法を身に付けることは、間接的な予防につながります。
生活リズムが整っている
統合失調症の発症のきっかけになるストレスは、睡眠不足や、生活リズムの乱れから生じるケースが少なくありません。
ライフイベントにおける目立つストレスばかりではなく、日常生活の中に鬱積していくストレスも見逃せない危険な要因です。規則正しい生活は、すべての病気に対する基本的な予防策です。
備えあれば憂い少なし
統合失調症には、ある種のなりやすい性格の傾向があり、あるいは病前性格のようなものが認められるというのは、確かなようです。
ただし、この性格が原因で発症するのか、それとも、性格そのものが統合失調症の前駆的症状と見るべきかについては、見解がわかれていますが、最近では前駆症状ではないかという説が優勢になってきているようです。つまり、統合失調症の特有の性格というのは、発症前のサインではないかということです。
この仮説にたてば、性格を変える努力をするよりも、統合失調症のリスクが高いということを自覚し、いざという場合に備えた対策を考えた方が有効だと言えます。つまり、早期治療に取り組む用意もしておくということです。
なぜ、それが有効かと言えば、精神科の医療技術の進歩で、統合失調症は、適切に早期治療に取り組めば、症状も治まり、再び社会復帰ができるようになってきたからです。のサインではないかということです。
ドパミン神経系には4つの経路がある
中枢においてドパミン神経系には4つの経路があります。
①中脳辺縁系神経路
②中脳皮質系神経路
③黒質線条体系神経路
④漏斗下垂体系神経路。漏斗下垂体系神経路というのは視床下部にある下垂体の漏斗系神経路のことです。視床下部にある下垂体はさらに,上に向かってのびている隆起部(隆起葉),下垂体の前部を占める前葉(主葉),前葉の後部に位置する中間部(中葉)の3部に分けられる。神経下垂体は腺下垂体の後部にあり,漏斗と後葉(神経葉)からなる。なお漏斗は正中隆起と漏斗茎に分けられる。乳汁分泌の作用があるホルモンであるプロラクチンを産生します。漏斗下垂体系にも働く抗精神病薬はプロラクチンホルモンの分泌に関係する漏斗下垂体系に作用することで乳汁分泌や月経障害、性機能障害などの副作用が現れることもあります。
神経細胞は脳の中に似たような作用を持つ神経細胞体の群れを作ってその神経が作る神経伝達物質の働きに対応した脳の他の部位に神経線維である軸索を送っています。これを神経を投射すると言います。例えばドパミン神経系といわれるA1~A17群は脳幹に細胞体を黒質や中脳腹側被蓋野に持っており神経線維を大脳辺縁系の線条核や大脳基底核の側坐核や前頭葉などに送りさらに視床下部にもドパミン神経系の神経細胞体を持っており下垂体は神経線維(軸索)を送っているのです。 因みに側坐核は運動系の中枢の大脳基底核に属する最も重要な神経核です。心の牽引車である前頭前野のすぐ後ろの奥にあり、ふたり以上の人間の心を連結するための連結回路(インターフェイス)とも言ってもいいのです。しかも一人の人間の前頭前野(前頭連合野)、大脳辺縁系、大脳基底核と緊密な神経連絡をしています。そして人間の感情、意欲の維持に極めて深く関与しています。
何故ストレスがかかるとステロイドやドパミン以外にノルアドレナリンという神経伝達物質が出るのでしょうか?ストレスがかかると不安や恐怖を感じたりします。ひどくなると鬱状態になることもあります。そのような心配や怖れを感知したノルアドレナリン作動性神経の活動性が高まり不安や怖れを感じる精神をさらに高めて他の神経にもストレスに対して準備させ対抗させようとするのです。s
それぞれ
①統合失調症の陽性症状
②統合失調症の陰性症状、認知機能
③錐体外路症状、パーキンソン病
④血漿プロラクチン値
に影響していると言われています。
表にまとめます。
| ドパミン神経系 | 影響 |
| 中脳辺縁系 | 統合失調症の陽性症状 |
| 中脳皮質系 | 統合失調症の陰性症状、認知機能 |
| 黒質線条体系 | 錐体外路症状、パーキンソン病 |
| 漏斗下垂体系 | 血漿プロラクチン値 |
統合失調症の原因はドパミンを増やす原因と同じです。ステロイドホルモンを出しやすい男女とも常にストレスを感じる人は常に快楽ホルモンであるドパミンを脳幹の中脳のA8、A9、10Aドパミン作動性神経群で出し続けることになります。A8は中脳辺縁系に過剰に投射してドパミンを伝えるのみならずA10とともに大脳辺縁系にもドパミンを流し込みます。A9は黒質緻密部からさらに被殻と尾状核の線条体に投射してドパミンを流し込みます。このA9は黒質線条体系に脳全体の80%のドパミンを大量に周辺の神経細胞に流しこむことによってストレスに耐えようとするのです。A10は大脳辺縁系の情動回路に陽性症状を引き起こすと大脳皮質に大量のドパミンを流して陰性症状を引き起こすのです。
本来ドパミン(ドパミン、dopamine)の作用は 運動の調節、ホルモンや循環の調整、学習、意欲、喜び、快楽に関与しているのだが過剰になると陽性症状と陰性症状と認知症が伴う統合失調症となってしまうのです。
現代の脳学者はパーキンソンと統合失調症を同じ原因で考えようとするので混乱を招いているのです。 パーキンソンは黒質の細胞にherpesが感染して細胞崩壊と脱落が生じて黒質線条体の機能障害でパーキンソンが起こったのです。一方、A8,A9,A10ドパミン作動細胞群が産生するドパミンがストレスのために増えたステロイドホルモンが異常に増やしすぎたために生じたのが統合失調症であるのです。
ドパミン神経細胞群(神経細胞集団)のA8細胞集団~A15細胞集団のそれぞれの役割を説明します。ついでにノルアドレナリン神経細胞群のA1細胞集団~A7細胞集団のそれぞれの役割も説明しておきます。
A1~A7細胞群はノルアドレナリンを分泌します。主に怒りや記憶や、覚醒を司ります。脳幹にあるA6細胞群は青いので青斑核と言われるのですがA6細胞群で怒りを引き起こすノルアドレナリンを最も多く分泌するので「怒りの中枢」とも言われます。因みにC神経細胞群から分泌されるアドレナリンは恐怖の感情を引き起こします。
A8細胞群:赤核後方部 (RRF) から 中脳辺縁系に投射する。A10細胞群と共に大脳辺縁系に投射し、中脳辺縁系を構成する。
A9細胞群:黒質・緻密部(SNc)から 線条体(尾状核と被穀)へ投射する。黒質線条体系を構成する。黒質線条体系は脳全体のドパミン量の80%を占める。黒質線条体系は錐体外路系の一部で、この部位のD2受容体遮断に関連し運動系が支障を起こしパーキンソン病のような錐体外路症状を起こすと言われますが実はパーキンソンは黒質にherpesが感染して黒質のドパミンを産生する細胞が崩壊して脱落しまってドパミンが作れなくなったからです。黒質線条体系は脳全体のドパミン量の80%を占めるのでその中で黒質の神経細胞体の働きがなくなると錐体外路症状が出現して錐体外路が正常に働かなくなってしまうからです。
セロトニン神経系はB神経系と言われます。セロトニン5-HT神経系(B系神経)は、左右対にB1〜B9まである。脳幹にある縫線核群の細胞の多くのものがセロトニンを含むがすべての縫線核群の細胞がセロトニンを含んでいるわけではないのです。
セロトニンの役割はカテコール・アミンの覚醒ホルモンの過剰分泌を抑制し、コントロールするのである。例えば、睡眠を誘い、活動を適度に制するのである。セロトニン( serotonin)は、別名5-ヒドロキシトリプタミン(英: 5-hydroxytryptamine、略称: 5-HT)は、必須アミノ酸トリプトファンから生合成される脳内の神経伝達物質のひとつで、動植物に広く分布する生理活性アミン、インドールアミンの一種です。5-HT神経系は、ノルアドレナリン神経系やドパミン神経系などとともに脳内モノアミン神経系である。
5-HT神経系は、脳幹の縫線核群に小さな集団として分布する5-HT神経が、広範な脳領域に投射して、多彩な機能に影響を与えている。
5-HT神経が標的細胞に作用する様式は、volume transmissionとしての特性を示す。
組織学的には神経終末と標的細胞との距離が比較的広く、通常のシナプス形成を示さない。
5-HT神経系は、痛みやストレスなどの内外環境からの覚醒・ストレス刺激には影響されない。
脳内のパターン形成機構によるリズム性運動(歩行運動、咀嚼運動、呼吸運動、グルーミングなど)で興奮し、覚醒状態における種々な活動に適度な緊張(抗重力筋の緊張や交感神経の緊張など)を与える役割がある。
5-HT神経は、他の神経からの刺激がなくても、自律的に低頻度のインパルスを出している。
5-HT神経は、その活動様式の点でも特徴があり、睡眠-覚醒の状態に依存した活動変化を示す。
5-HT神経は覚醒時には規則的な(3-5Hz)発射活動があり、徐波睡眠に移行するとその活動が減弱、レム睡眠になると、完全に消失する。
5-HT神経の活動特性は、覚醒時に抵頻度発射を継続して、標的細胞のシナプス間隙に一定の5-HTを分泌させ、覚醒状態を維持する。
5-HT神経の発射頻度を決めるのは、オートレセプター(5HT1A受容体)によるネガティブフィードバックである。
5-HT1A受容体の数が多いと、自己抑制が強くなり、発射頻度が低く抑えられる。
覚醒時の5-HT神経系の活動が抑制された状態は、うつ病や慢性疲労症候群などの症状を惹起すると考えられる。
縫線核( raphe)について説明しましょう。
縫線核 rapheは、発生学的に最も古い脳である脳幹の中央、左右の脳が正中で縫い合わされる部位にある。
蛍光法によって、縫線核群の細胞の多くのものがセロトニンを含むことが知られている。
しかし縫線核のニューロンのすべてがセロトニン含有ニューロンではない。
縫線核の周囲には、歩行、咀嚼、呼吸などのリズム運動を形成する中枢が局在する。
縫線核群は尾側と吻側(背側)に大別される。
セロトニン含有ニューロンの軸索は非常に多くの側枝を出している。
尾側から下行線維、吻側から上行線維が出る。
尾側縫線核群:延髄腹側に分布する。不確縫線核、淡蒼縫線核、大縫線核に分けられる。
軸索は脊髄を下行する。各神経細胞に投射する。
吻側の縫線核群:中脳の背側に分布する。背側縫線核、正中縫線核などに分けられる。
軸索は、小脳、中脳、間脳、辺縁系、大脳皮質などに投射し、各種の生理機能に影響を与える。
ドパミンD2受容体は、ドパミンにより活性化されて興奮性の情報伝達を行うのですがドパミンが減ってしまったためにパーキンソンが起こるのです。ドパミンは、快楽の神経伝達物質であると同時に快楽をもたらす運動調節や脳の快楽にかかわる向上心や意欲や学習の達成感に関わる脳内の興奮性の神経伝達物質です。 脳内のドパミン量が不足するとパーキンソン病になり、過剰になると統合失調症になるだけでドパミンD2受容体の働きがパーキンソン病や統合失調症に全く関りがないのです。何故統合失調症になるかは最後までこの論文を読めばお判りになります。乞うご期待!!!
錐体路系や錐体外路系とか錐体外路障害とか錐体外路症状とは何でしょうか?
錐体路とは随意運動に関与するもので、その経路は前頭葉の運動野やから延髄の錐体交差で反対側に交差し、脊髄の前角に至るまでをいいます。この経路は、運動の「オン・オフ」のはたらきをし、錐体路の障害では運動麻痺まひがみられます。一方、錐体外路とは大脳深部の底部にある大脳基底核を中心とする複雑な経路のことです。運動や筋緊張を調整する働きをしています。従って錐体外路の障害では、筋緊張の異常や、さまざまな不随意運動がみられます。錐体外路症状の中で一番重症なのは癲癇です。癲癇では一切の筋肉が動かなくなるどころか意識も失ってしまうのです。つまり錐体外路症状はすとは大脳深部の底部にある大脳基底核に感染したherpesによって生じるのです。異常な運動症状を示すのは錐体外路症状と考えていいのです。
多くの種類の異常な運動症状がありますが、これらの異常は運動過少と運動過多の2種類に大別されます。運動過少を呈する症状は、固縮、無動などであり、パーキンソン病や、パーキンソン病に類似した症状を呈するパーキンソン症候群でしばしばみられる異常な運動は運動過少の症状である。 運動過多を呈する症状は、①振戦、②舞踏運動、③片側バリズム、④アテトーゼ、⑤ジストニアなどであり、不随意運動です。運動過多の振戦、運動過少の固縮と無動はパーキンソン病の三大徴候であります。①振戦とは自分が動かそうと思っていないにもかかわらず生じてしまう「ふるえ」のことです。 ②舞踏運動とは、手をチョコチョコと動かしたりすることが多く、不規則に繰り返される短くやや速い不随意運動を特徴とし、ダンスをしているように見えるので舞踏運動と言います。 ③片側バリズムとは 英語でヘミ(片側)バリスムとも言います。舞踏運動の一種で、片腕または片脚を投げ出すような激しい不随意運動です。 動きは舞踏運動より大きく激しくなります。 ④アテトーゼとはゆっくり流れるようにうねる連続的な不随意運動です。 これは通常、手と足に現れます。 ⑤ジストニアとは意志によらない自分では制御できない運動(不随意運動と呼びます)で、比較的長い筋肉の収縮により生じます。 ジストニアは体の様々な部位にみられ、斜頸、顔面痙攣、書痙がみられます。 書痙とは字を書く時に緊張して手が震え、うまく書けなくなってしまうことです。これらの錐体外路症状は、すべて大脳基底核の神経細胞に感染したヘルペスによるものなのです。大脳基底核に感染したヘルペスが大脳皮質―大脳基底核ループにまで異常な電気信号を送ると重症な癲癇になるのは『癲癇はなぜ起こるのか』で詳しく説明しました。
錐体外路症状と言うと、主に体性運動系の障害を指すのですが、その他の運動系や非運動系にも錐体外路症状は現れるので例えば、パーキンソン病における動作緩慢は眼球運動系にも認められ、それは癲癇はなぜ起こるのかで説明したように大脳皮質―大脳基底核ループの眼球運動ループがヘルペスによる障害された結果なのです。
パーキンソン病患者では、herpesが感染により黒質緻密部に感染して黒質緻密部のドパミン神経細胞の変成、脱落が生じて、線条体で放出されるドパミンが減少して、上に述べた無動、筋固縮、振戦などの運動症状が生じるのです。パーキンソン病に対するドパミン補充療法は、黒質線条体系を刺激してその運動症状を改善するが、同時にA10細胞群がある中脳の腹側被蓋野 (VTA)の中脳皮質辺縁系も刺激されるので、ドパミン作動薬による薬物依存・離脱症状、ドパミンの濃度が高くなり新たなる病気を作ってしまうのです。ドパミンが多くなりすぎると不快な身体症状、不眠、不安、振戦、発汗、痙攣発作、妄想、幻覚、体感幻覚までもが生じて薬物による統合失調症が生まれるのです。 体感幻覚(たいかんげんかく)とは手足や臓器などの奇妙な違和感を感じることです。「腸が腐る」「脳が溶けている」など独特の言い回しで表現され、統合失調症でみられます。因みに統合失調症の幻覚には幻聴、幻視、幻臭、体感幻覚などがあります。
A10細胞群:腹側被蓋野 (VTA)に二種類あり①大脳辺縁系(側坐核・海馬・扁桃体・嗅結節・中隔・梨状葉)の情動回路に投射するのと②大脳皮質系(前頭前野)の不安で活性化される回路に投射する二種類です。VTAは ventral tegmental area, ventral tegmentumの略語です。①の大脳辺縁系(側坐核・海馬・扁桃体・嗅結節・中隔・梨状葉)の情動回路は行動および情動(感情)に関与しており、統合失調症の陽性症状を生み出します。 側坐核とはherpesが感染すると癲癇を起こす運動系の中枢の大脳基底核に属する最も重要な神経核です。心を支えてくれる牽引車である前頭前野のすぐ後ろの奥にあり、人間のよこしまな 本能(大脳辺縁系)と 理性(大脳皮質の前頭前野)を結び付けるインターフェイス(接続装置)で、 そして前頭前野(前頭連合野)、大脳辺縁系、大脳基底核と緊密な神経連絡をして人間の本能であるエゴイズムを制御しています。 前頭前野(前頭連合野)とはヒトの大脳で感覚野,運動野には属さない部位を連合野association areaとよぶ。前方の前頭葉frontal lobeに位置する連合野が前頭連合野とよばれる。この部位は前頭前野,あるいは前頭前皮質とよばれることもある。知能指数(IQ)のレベルと最も関連する脳部位は、「前頭前野」です。前頭前野の皮質の成長パターンがIQレベルによって異なるのです。前頭前野は「脳の中の脳」と言われていますから「理性の座」そのものです。
知能指数を生み出す脳の部位はどこにあると思いますか?知能(IQ)レベルと最も関連する脳部位は、「前頭前野」と呼ばれる場所です。知能は知的能力の略語です。「前頭前野」の 同義語には前頭連合野、前頭前皮質、前頭顆粒皮質の三つがあります。前頭前野の皮質の成長パターンがIQレベルによって異なるので知能を上げるには、つまり大脳皮質の成長を刺激するためには毎日知的能力を磨く努力をすることが一番大切です。
前頭前野はヒトをヒトたらしめ,思考や創造性などの知性を担う脳の最高中枢であるのです。前頭前野(前頭連合野、前頭前皮質、前頭顆粒皮質)は系統発生的にヒトで最もよく発達した脳部位であるとともに,個体発生的には最も遅く成熟する脳部位である。一方老化に伴って最も早く機能低下が起こる部位の一つでもある。この脳部位はワーキングメモリ、反応抑制、行動の切り替え、プラニング、推論などの認知・実行機能を担っている。 ワーキングメモリとは脳の機能の一つで、情報を一時的に記憶・整理することです。前頭前野(前頭連合野、前頭前皮質、前頭顆粒皮質)はまた、高次な情動・動機づけ機能とそれに基づく意思決定過程も担っている。さらに社会的行動、葛藤の解決や報酬に基づく選択など、多様な機能に関係しています。
嗅結節とは嗅覚の一次中枢である嗅球から直接投射を受ける嗅皮質領域の一部であり、同時に側坐核とともに腹側線条体を構成する脳領野である。大脳の嗅結節と呼ばれる部位に、食べ物の匂いで「食べたい」(摂食)モチベーションを引き起こす神経回路と、危険な匂いにより「避けたい」(警戒・忌避)モチベーションを引き起こす神経回路が、別々に分かれて存在します。動物にとっては長い進化の中で「摂食」と「警戒」は臭いの感覚によってなされたので動物の中で霊長目の長であるので人間も今なお嗅覚の神経路を保存維持しているのです。
中隔核(中隔)とは大脳辺縁系に属し,海馬や扁桃体と密接な関係にあり,視床下部や脳幹とも神経連絡があることなどから, 自律神経の上位中枢として働いている。中隔核は喜怒哀楽を司る偏桃体の「楽」の感情と深い関りがあるので「楽しい感覚」に関与する神経核であり、中隔核(中隔)が破壊されると「楽」を感ずる情感が無くなり怒りに対する抑制がなくなってしまい怒り狂うのです。
中隔核には脳幹の中脳にあるセロトニン作動性ニューロンである縫線核からの入力があり,自律神経機能や情動の形成に深く関わっています。
縫線核とは中脳から脳幹の内側部に分布する細胞集団で、9つの神経核B1~B9よりなりこれをB系神経とかセロトニン5-HT神経系と言い、左右にB1〜B9まである。セロトニン作動性神経細胞の核です。大きく ①吻側核群、②背側縫線核、③尾側核群の三つに細分類されそれぞれ働きが異なるのでセロトニン作動性神経細胞の核は多彩な働きを持っており縫線核の投射は脳全体にわたっています。セロトニンは受容体だけでもサブタイプを含めて14種類あるため、 セロトニンの作用は非常に多様・複雑、そしてわかりづらいです。セロトニンはどんな物質?とは一言で言えないのです。睡眠覚醒・歩行・呼吸などのパターン的な運動や注意・報酬などの情動や認知機能にも関与しています。 セロトニンの産生はアミノ酸のトリプトファンからトリプトファン水酸化酵素によりまず5-水酸化トリプトファンになった後に5-水酸化トリプトファン脱炭酸酵素によりセロトニンになる。脳内で水酸化酵素を持つのは縫線核群の5-HT神経系の細胞か松果体のメラトニン産生細胞だけです。 セロトニンの分泌亢進因子は歩行、咀嚼、呼吸のリズム運動と太陽光です。 セロトニンの分泌抑制因子は慢性ストレスで、ストレスによって活性化される神経である視床下部室傍核のCRH(corticotropin releasing hormone訳は副腎皮質刺激ホルモン放出ホルモン)が縫線核に投射して5-HT神経の活動を抑制するのです。一過性のストレスでは5-HT神経の活動を抑制しません。何故でしょうか?今は分かりませんがいずれ答えを出します。
大脳皮質下領域とは大脳の皮質でもなく、しかも髄質でもない大脳の深部にある領域で、大脳辺縁系の海馬、扁桃体や、間脳の視床や大脳基底核の側坐核、尾状核、被殻、淡蒼球などが含まれ、系統発生的に古いとされる領域で、 運動機能や記憶・情動・意欲などに関与するとされています。大脳皮質下領域は曖昧な言葉です。
梨状葉皮質はその梨状葉ニューロンのニオイ分子に対する反応が嗅球の僧帽細胞の反応と似ており梨状葉皮質が行っている役割は,ニオイの識別です。
腹側線条体とは,側坐核を中心に,嗅結節および尾状核・被核の最も腹側部を含む領域で,その解剖学的な位置から辺縁系と運動系のインターフェース(接点)の役割も果たしています。更に動機づけに最も深く関わっている嗅結節の内側部が薬物依存の発症に関わるのは、ドパミンは快楽をもたらす神経伝達物質だから今なお快楽こそ動機づけの人間の行動の原点になっているのです。
大脳辺縁系とは人間の大脳の辺縁にある神経系で情動の表出、食欲、性欲、睡眠欲、意欲、などの本能、喜怒哀楽、情緒、神秘的な感覚、睡眠などの本能と情動を司っている神経系ですが一部は本能の記憶や自律神経活動にも関与していますが基本的には「本能の座」であるので高等な大脳皮質とは直接かかわりはないのです。
A10のドパミン神経の細胞群の話に戻りましょう。 A10のドパミン神経の細胞群は中脳の腹側被蓋野が正式の名称です。「被蓋」という意味は「中脳の中身をすっかりおおい隠すように作られた、縁(ふち)のある蓋(ふた)」です。腹側被蓋野(ふくそくひがいや、ventral tegmental area, ventral tegmentum、VTA)は哺乳類の脳における中脳の一領域であり、被蓋腹側に位置する。 被蓋とは脳幹の背側の領域を広く指す言葉であり、系統発生的に古い部分であり赤核や黒質も被蓋に含まれるのです。 赤核とは赤核は中脳被蓋部腹側,動眼神経核の側方に位置する赤色の核ですが,その機能については十分には理解されていない。統合失調症の原因を突き止めるためには A10のドパミン神経の細胞群を十分理解する必要があるのです。何故ならばA10のドパミン神経の細胞群が産生する大量のドパミンこそが統合失調症の原因だからなのです。何故A10のドパミン神経の細胞群が大量にドパミンを産生するのかを明らかにするのがこの論文の目的であるのです。
この腹側被蓋野からは二種類のドパミン神経路が出ています。ひとつは大脳辺縁系へ投射する中脳大脳辺縁系路であり二つ目は前頭葉に投射する中脳大脳皮質路の二つです。中脳大脳辺縁系は統合失調症になりやすい人は情動(感情)が極めて敏感でかつ動かされやすくすぐに興奮してしまうとこの中脳大脳辺縁系路が刺激され活性化されてしまうのです。この神経路の異常が統合失調症の陽性症状に関わっていると言われています。一方、②中脳大脳皮質系路は統合失調症になりやすい人は少しのストレスや不安などの負の要因により刺激され中脳大脳皮質系路はすぐに活性化してしまうのです。この経路の異常が陰性の症状が発現するのです。しかし現代世界には多かれ少なかれ感性が敏感な人や不安を感じやすい真面目な人ばかりです。僕もその一人ですが鬱になったことはいくらでもありますが統合失調症ではありません。しかも私はherpes脳炎である大脳辺縁系の海馬炎と偏桃体炎にすでになっていますが残念ですが統合失調症ではありません。というのは統合失調症を自分自身が体験すればもっとこの病気を理解できるからです。
① の中脳大脳辺縁系路正常な神経路の働きは例えば快楽を得るために快楽の情動回路であるA10細胞群のドパミン作動神経はどのようにして活性化されるのでしょうか?例えば「さすってもらったら痛みが収まった」経験が快情報として快楽の中枢である扁桃体が認識して記憶され隣接する海馬体へ格納記憶され、更に、この快の記憶を再び実現したいという情動を支配する扁桃体がA10を活性化します。 A10活性化によりドパミンが出され側坐核を興奮させて、視床内背側核 → 前頭前野 → Yakovlevの情動回路を作動させて、快楽の情動行動を起こす方向へと進むのは正常な人の話です。Yakovlevの情動回路は扁桃体→視床背内側核→前頭葉眼窩皮質後方→側頭葉前方→扁桃体に戻ります。この正常なドパミン作動性中脳大脳辺縁系路が異常になって統合失調症の陽性症状が出るのですが陽性症状という異常を起こす原因を世界中の医者は知らないので私が答えを出そうとしているのです。ただ現在の統合失調症の効く薬はすべてがドパミンを減らす薬ではなくA10細胞群のドパミン作動性神経からシナプスに出されたドパミンを受け取る回路の神経のドパミン受容体を不活化させるものばかりですからこのドパミン受容体とherpesとの問題ではないかと考えています。そのためにドパミン受容体のすべてを勉強します。答えは必ず出しますから乞うご期待を!!!Yakovlevの情動回路は扁桃体→視床背内側核→前頭葉眼窩皮質後方→側頭葉前方→扁桃体。Yakovlevの情動回路のゴロの覚え方は、変死前に即答返答やれ。
②の中脳大脳皮質系路の正常な働きの例として不安で活性化される皮質系(前頭前野)回路は、不安やストレスにより活性化し、時に負の行動を起こす統合失調症の陰性症状状を発現させると言われています。又、ストレス刺激がA10経路を活性化すると、前頭前野のドパミンD1受容体を介して前頭前野の行う認知・思考・ワーキングメモリ等の機能をも抑制してしまうのですがこれも正常な人で見られることがあるのです。
中脳の腹側被蓋野(ふくそくひがいや、ventral tegmental area, ventral tegmentum、VTA)は哺乳類の脳における中脳の一領域であり、中脳被蓋腹側に位置する。被蓋とは脳幹の背側の領域を広く指す言葉であり、系統発生的に古い部分である(赤核や黒質も被蓋に含まれる)。この中の腹側被蓋野は黒質や赤核に囲まれた内側の領域である。赤核は中脳被蓋部腹側,動眼神経核の側方に位置する赤色の核で中脳被蓋部において大きな部分を占める核である。赤核の名前の由来は鉄分を多く含みピンク色を呈するからです。卵円形の神経核。大脳皮質や小脳から情報の入力を受け、不随意運動の調節を行う。
中脳腹側被蓋(VTA)のドパミン神経は報酬や目標志向型の行動に中心的な役割を担っている。VTAのドパミン放出神経細胞は様々な入出力パターンを持つものが混在しており、中にはドパミンに加えてGABAやグルタミン酸を放出するドパミン神経もいる。動機付け信号はVTAのドパミン神経だけでなく、独自の回路を持っている非ドパミン神経によっても生じている。ドパミン神経も非ドパミン神経も局所の抑制的、興奮的回路を使って入力された情報を統合し、出力発火パターンを形成している。様々な個々の入力、出力、局所回路は報酬行動や忌避行動を引き起こすのに十分であり、この小さな細胞集団の行動への寄与は非常に重要である。
中脳の腹側被蓋野(ふくそくひがいや、ventral tegmental area, ventral tegmentum、VTA)のドパミン神経の重要なポイントをまとめましょう。
VTAのドパミン神経は様々な動機付け行動に寄与している。異なった行動は独自の神経回路を持った異なったドパミン神経によって媒介されている。VTAの出力神経細胞は他の脳領野からの情報を統合するだけでなく、GABA神経やグルタミン酸神経からの情報も統合し、微小回路を形成している。VTAのGABA神経やグルタミン酸神経の一部はドパミン神経と同じ脳領野から入出力を受けている。VTAの神経細胞の一部は、ドパミンとともに、グルタミン酸もしくはGABAを共放出する。
VTAのドパミン神経はドパミン産生酵素であるチロシン水酸化酵素を発現し、ドパミンを放出する神経細胞と定義されます。VTAのドパミン神経は正や負の強化学習、意思決定、作業記憶、報酬の顕著性、刺激の顕著性、忌避刺激において重要な役割を担っている。これらの行動上の混在性は、VTAのドパミン神経が様々な発火パターンや入出力パターンを持っていることを反映している。VTAのドパミン神経の発火パターンは、局所性のグルタミン酸神経入力、GABA神経入力のみならず、様々な脳領野からの入力によって制御されている。VTAのGABA神経やグルタミン酸神経は局所回路の形成に関与するのみならず様々な脳領野に長距離投射を行っている。
A11ドパミン細胞群(神経核)からA15ドパミン細胞群(神経核)までの神経核は中脳ではなく間脳に存在しています。間脳は視床と視床下部から成り立っています。
A11細胞群:間脳後部 から A13、 A14細胞群と共に視床下部、脊髄側角に投射し、視床下部脊髄路を構成する脊髄後角/三叉神経脊髄路核の侵害受容神経細胞に対する抑制に関与しており、片頭痛を制御していると示唆されています。A11細胞群機能の低下により、レストレスレッグス症候群、下肢静止不能症候群、むずむず足症候群の症状が起こる。出現症状は、下肢を中心に不快な異常感覚が、特に夕方から深夜にかけて出現。「むずむずする」「じっとしていられない」「痒い」「ピンでなぞられているような」「針で刺すような」「火照るような」「蟻やミミズなどの虫が這っているような」などの異様な感覚が現われ、時には「振動」を感じる場合もある。
A12細胞群:視床下部・弓状核 から 下垂体正中隆起、下垂体中間葉、下垂体後葉に投射。下垂体の隆起漏斗ドパミン作動系を構成しており下垂体ホルモン調整を行っている。
A13細胞群:A11細胞群の一部と共に視床下部に投射し、不確帯視床下部ドパミン作動系を構成する。不確帯(ふかくたい zona incerta)とは腹側視床の一部を成す、哺乳類の脳の灰白質の小領域。 内包の内側に位置し、狭義の視床(背側視床)の腹側に位置する。 視床網様核と隣り合わせの位置である。
A14細胞群:A11細胞群の一部と共に視床下部に投射し、不確帯視床下部ドパミン作動系を構成する。
A15細胞群:視床下部のドパミン細胞群。
A16細胞群は嗅球にあります。
A16細胞群:嗅球のドパミン細胞群。嗅球はヒトにおいては前頭葉の下に位置する長円形の小さな構造である。鼻腔中の嗅上皮で匂い受容を行う嗅神経細胞からの入力を受け、嗅皮質に出力する。嗅球内にはドパミン合成酵素であるチロシン水酸化酵素(TH)を発現している介在ニューロンが多数存在し、ドパミンの生合成が行われている。さらに、ドパミン代謝酵素の一つである、芳香族 L-アミノ酸脱炭素酵素(AADC)の発現が見られることより、ドパミンの合成がおこなわれているのです。
A17細胞群:網膜のドパミン細胞群。人の網膜において、ドパミン放出型アマクリン細胞 (DA細胞) は光照射刺激によりドパミンを放出する。そのドパミンは傍分泌により様々な種類の網膜細胞に作用し、結果として明順応信号として機能している。傍分泌(ぼうぶんぴ・ぼうぶんぴつ、 Paracrine signaling)とは、細胞間におけるシグナル伝達のひとつで特定の細胞から分泌される物質が、血液中を通らず組織液などを介してその細胞の周辺で局所的な作用を発揮することである。
ドパミン受容体は、D1、D2、D3、D4、D5受容体あります。ドパミンが受容体と結合すると神経細胞に電位変化が起こったり、細胞内の情報伝達系が動いてさまざまな変化を受け手の神経細胞に引き起こします。役目を終えたドパミンはまた神経末端のシナプスにあるドパミントランスポーターに、たんぱく質として取り込まれて再利用される仕組みになっています。
受容体は、興奮性、抑制性と2つに分類されていますが、まだ不明点が多く単純ではなさそうです。 ①興奮性を担う受容体:D1、D5受容体。 ②抑制性を担う受容体:D2、D3、D4受容体。例えば前頭葉のNMDA受容体やドパミン受容体の異常によってドパミンの分泌が亢進し、D2受容体を刺激してしまうため、視床が興奮して、知覚過敏の状態になる錐体外路症状を生ずる。また、D2受容体が長期に遮断されると、遅発性ジスキネジアなどの運動障害を引き起こす。このD2受容体は②抑制性を担う受容体に分類されているのに何故視床が興奮して、知覚過敏の状態になる錐体外路症状を生ずるのでしょうか?また、D2受容体が長期に遮断されると、遅発性ジスキネジアなどの運動障害を引き起こす。このD2受容体は②抑制性を担う受容体に分類されているのに何故抑制を遮断すれば興奮して遅発性ジスキネジアなどの運動障害を引き起こすことはないはずですのに?
それぞれの受容体の働きを見ましょう。
D1受容体:中枢神経系で最も豊富なドパミン受容体で神経細胞の成長を調節。線状体背側部に発現が見られ 、黒質に投射する。腎血管や腸間膜動脈にも存在し、血管拡張をもたらす。そのほかいくつかの行動反応に関係。
D2受容体:筋緊張異常や統合失調症に関連していると考えられています。線状体背側部に発現が見られ淡蒼球に投射する。ドパミン作動性神経細胞や軸索終末部にも存在し、ドパミンの産生制御と放出制御を行う。ドパミン神経系のA12細胞群のドパミンは、下垂体前葉のプロラクチン産生の細胞のD2受容体に作用し、プロラクチン分泌を抑制する。
D3受容体:大脳辺縁系に局在、認知的および感情的な機能に働くことを示し、統合失調症やパーキンソン病の治療薬の標的です。
D4受容体:認知や情動と関連している大脳皮質や辺縁系に多く存在。統合失調症やパーキンソン病の治療薬の標的。行動表現型に関連していると考えられています。統合失調症患者さんの死後脳の線条体ではD4受容体が著しく増加していることを示された。
D5受容体:脳の辺縁領域の神経細胞で見られ、D1受容体よりも10倍高いドパミン親和性をもつのが特徴。
パーキンソン病では、黒質 → 線条体(背側)の神経細胞の変性により線条体のドパミンの枯渇により発生と言われています。また、ドパミン トランスポーターの顕著に低下がみられます。ドパミントランスポーターは、神経伝達物質ドパミンをシナプス間隙から細胞質ゾルにポンプで戻す膜貫通タンパク質です。更に細胞質ゾルでは、他のトランスポーターがドパミンを小胞に隔離して貯蔵し、後で放出します。
すでに述べましたが、パーキンソン病は、運動障害、非運動障害も来たす病気で、非運動障害の中には、神経精神症状(うつ、幻覚、認知症など)、睡眠症状 (むずむず足症候群、REM(レム)睡眠関連行動異常(寝言、夜驚症など、日中過眠など)、感覚症状 (痛み、締め付け感)。
パーキンソン病の自律神経障害には骨盤臓器障害 (過活動膀胱・便秘)、流涎、嚥下障害、胃部不快、起立性低血圧、四肢の冷え、網状皮斑、発汗異常、脂漏性顔貌、体重減少などが含まれます。過活動膀胱とは、急に尿意をもよおして、何回もトイレに行ったり、トイレに間に合わず尿をもらしてしまったりする病気のことをいいます。 過活動膀胱になると、尿が十分にたまらないうちに膀胱の筋肉が勝手に収縮して、尿を出そうとします。網状皮斑とは、皮膚に赤色や紫色の網目模様がみられる状態を指します。「リベドー」と呼ばれます。皮膚に分泌する動脈・毛細血管・静脈の調整機構がうまくいかないために、血液の循環が悪くなることを原因として発症します。パーキンソン病も他のすべての症状の原因がヘルペスであるように過活動性膀胱も網状皮斑もすべてヘルペスが原因です。
ドパミンやノルアドレナリンやアドレナリンの三つの神経伝達物質を放出する脳内の神経細胞集団(神経核)起始部のある部位をまとめておきましょう。
ドパミン神経系は、A8~A17でありすでに説明しました。
A8~A10神経群は中脳に位置し、A11~A15神経群は、間脳に位置します。
A8~A10神経群は中脳に位置し、A11~A15神経群は、間脳に位置します。
ノルアドレナリン神経系の神経細胞集団は、A1~A3は、延髄にありA4~A7は、橋に位置します。
一方、アドレナリン神経系の神経細胞集団(神経核)はC1~C3で中枢神経系におけるアドレナリン作動性の神経細胞は、すべて延髄にあり主に次の三つの延髄の部位にある。
C1:延髄の腹外側にありノルアドレナリン作動性神経細胞核A1に近接する。尾側の細胞群は、視床下部に上行性投射をし、循環器系や内分泌系の調節を行う。吻側の細胞群は、脊髄に下行性投射をし、交感神経の節前線維を形成する。
C2:延髄の背側にありノルアドレナリン作動性神経細胞核A2と一部重なる。C1、C2共に視床下部の室傍核に上行性投射をし、循環器系や内分泌系の調節を行う。
C3:延髄の吻側正中線近傍に位置し、視床下部、青斑核などに上行性投射、脊髄に下降性投射を行う。
アドレナリンの神経核は延髄に存在しているが、実はアドレナリン神経ではなく、ノルアドレナリン神経として副腎髄質へ投射して、ノルアドレナリンの刺激で副腎髄質からのアドレナリンが分泌される。だからといってノルアドレナリンの神経核と名付けると作用の違うノルアドレナリンの神経核が二つも存在ししかも機能が異なるのでアドレナリンの神経核はノルアドレナリンの刺激ではあるのですが最終的には副腎髄質からのアドレナリンが分泌されるのでアドレナリンの神経核という名前になるのです。
そもそも、アドレナリンを分泌する副腎髄質、つまり副腎は腎臓の隣にあるため、中枢には存在していない。一方、ノルアドレナリンを分泌する青斑核を中心とした神経核は全て中枢に存在している。
| 神経集団番号 | 神経集団名称 | 主な作用 |
| C1 | 下オリーブ核・外側網様体の間 | 副腎髄質からのアドレナリン分泌作用 |
| C2 | 孤束核 | |
| C3 | 背側縫線核 |
ついでに言えば末梢神経系の節後神経細胞は、ノルアドレナリンと共にアドレナリン作動性でもある。脊髄中の節前神経細胞よりアセチルコリン性の入力を受け、ノルアドレナリン性の出力を内臓器官に与える。その結果、血管の収縮、血圧の上昇、心拍数の増加、などを引き起こす。
前頭葉のNMDA受容体やドパミン受容体の異常によってドパミンの分泌が亢進し、D2受容体を刺激してしまうため、視床が興奮して、知覚過敏の状態になる錐体外路症状を生ずる。また、D2受容体が長期に遮断されると、遅発性ジスキネジア などの運動障害を引き起こす。
パーキンソン病患者では、黒質・緻密部にあるドパミン神経細胞の変成、脱落により、 線条体で 放出されるドパミンが減少して、無動、筋固縮、振戦などの運動症状が生じる。
パーキンソン病に対するドパミン補充療法は、黒質線条体系を刺激してその運動症状を改善するが、同時に中脳皮質辺縁系も刺激されるので、ドパミン作動薬による薬物依存・離脱症状、薬物の血中濃度の高低変化で出現する不快な身体症状、不眠、不安、振戦、発汗、痙攣発作、妄想、幻覚、体感幻覚)が生じる可能性がある。一部はA10細胞群と共に前頭前皮質に投射し、中脳皮質系を構成する。
A10細胞群:
①腹側被蓋野 (VTA) → 辺縁系(側坐核・海馬・扁桃体・嗅結節・中隔・梨状葉)の情動回路と
②腹側被蓋野 (VTA) → 皮質系(前頭前野)不安で活性化される回路に投射。
行動および情動(感情)に関与すると言われ、統合失調症の陽性症状との関連も示唆されている。
① 快の情動回路の作動を例えると「さすってもらったら痛みが収まった」快情報として扁桃体が認識 → 記憶 として海馬へ格納。この快の記憶が原因で扁桃体がA10を活性化 → A10活性化によるドパミンが側坐核を興奮させて、視床内背側核 → 前頭前野 → Yakovlevの情動回路を作動させて、快の情動行動を起こす方向へと進む。
② 皮質系(前頭前野)不安で活性化される回路は、不安やストレスにより活性化し、負の行動を起こす統合失調症の陰性症状の発現。ストレス刺激がA10経路を活性化すると、前頭前野のドパミンD1受容体を介して前頭前野の行う認知・思考・ワーキングメモリ等の機能を抑制。
A11細胞群:
間脳後部 → A13、 A14細胞群と共にに視床下部、脊髄側角に投射し、視床下部脊髄路を構成する脊髄後角/三叉神経脊髄路核の侵害受容神経細胞に対する抑制に関与。片頭痛を制御していとの示唆れている。
A11細胞群機能の低下により、レストレスレッグス症候群、下肢静止不能症候群、むずむず足症候群の症状が起こる。出現症状は、下肢を中心に不快な異常感覚が、特に夕方から深夜にかけて出現。「むずむずする」「じっとしていられない」「痒い」「ピンでなぞられているような」「針で刺すような」「火照るような」「蟻やミミズなどの虫が這っているような」などの異様な感覚が現われ、時には「振動」を感じる場合もある。
A12細胞群:
視床下部・弓状核 → 正中隆起、下垂体中間葉、下垂体後葉に投射。隆起漏斗ドパミン作動系を構成する。
下垂体ホルモン調整を行っている。
A13細胞群:
A11細胞群の一部と共に視床下部に投射し、不確帯視床下部ドパミン作動系を構成する不確帯(zona incerta, ZI)。
A14細胞群:
A11細胞群の一部と共に視床下部に投射し、不確帯視床下部ドパミン作動系を構成する。
A15細胞群:
視床下部のドパミン細胞群。
A16細胞群:
嗅球のドパミン細胞群。
A17細胞群:
網膜のドパミン細胞群。
ドパミン受容体は、D1、D2、D3、D4、D5受容体と5種類あるとわかっています。
ドパミンが受容体と結合すると神経細胞に電位変化が起こったり、細胞内の情報伝達系が動いてさまざまな変化を受け手の神経細胞に引き起こします。役目を終えたドパミンはまた神経末端のシナプスにあるドパミントランスポーターに、たんぱく質として取り込まれて再利用される仕組みになっています。
受容体は、興奮性、抑制性と2つに分類されていますが、まだ不明点が多く単純ではなさそうです。
興奮性を担う受容体:D1、D5受容体。
抑制性を担う受容体:D2、D3、D4受容体。
D1受容体:
中枢神経系で最も豊富なドパミン受容体で神経細胞の成長を調節。線状体背側部に発現が見られ 、黒質に投射する。腎血管や腸間膜動脈にも存在し、血管拡張をもたらす。そのほかいくつかの行動反応に関係。
D2受容体:
筋緊張異常や統合失調症に関連していると考えられています。線状体背側部に発現が見られ淡蒼球に投射する。ドパミン作動性神経細胞や軸索終末部にも存在し、ドパミンの産生・放出制御を行う。ドパミン神経系のA12細胞群のドパミンは、下垂体前葉のプロラクチン産生の細胞のD2受容体に作用し、プロラクチン分泌を抑制する。
D3受容体:
大脳辺縁系に局在、認知的および感情的な機能に働くことを示し、統合失調症やパーキンソン病の治療薬の標的です。
D4受容体:
認知や情動と関連している大脳皮質や辺縁系に多く存在。統合失調症やパーキンソン病の治療薬の標的。行動表現型に関連していると考えられています。
統合失調症死後脳の線条体ではD4受容体が著しく増加していることを示された。
D5受容体:
脳の辺縁領域の神経細胞で見られ、D1受容体よりも10倍高いドパミン親和性をもつのが特徴。
パーキンソン病では、黒質 → 線条体(背側)の神経細胞の変性により線条体のドパミンの枯渇により発生と言われています。また、ドパミン トランスポーターの顕著に低下がみられます。
パーキンソン病は、運動障害、非運動障害も来たす病気で、非運動障害の中には、神経精神症状(うつ、幻覚、認知症など)、睡眠症状 (むずむず足症候群、REM(レム)睡眠関連行動異常(寝言、夜驚症など、日中過眠など)、感覚症状 (痛み、締め付け感)。
自律神経障害には骨盤臓器障害 (過活動膀胱・便秘)、流涎、嚥下障害、胃部不快、起立性低血圧、四肢の冷え、網状皮斑、発汗異常、脂漏性顔貌、体重減少などが含まれます。
それでは中脳のドパミン神経核の神経集団名A8細胞群~A10細胞群の三つの神経集団の正式な名称と主なドパミン神経系の作用を見ましょう。A9細胞群とA10細胞群が極めて重要な作用を持っています。作用についてはできる限り詳しく説明しましょう。細胞集団を細胞群と言います。
A8の正式名称は赤核後核とも赤核後方部(RRF)とも言います。作用はA10細胞群と共に中脳辺縁系を構成して大脳辺縁系に投射する。
A9の正式名称は黒質緻密部です。作用は新線条体へ投射する。黒質線条体系を構成する。黒質線条体系に脳全体のドパミンの量の80%を占める。
パーキンソン病の関連部位として錐体外路運動に関与。新線条体は線条体(しんせんじょうたい neostriatum)とも、背側線条体とも呼ばれ、被殻(ひかく putamen)と尾状核(びじょうかく caudate nucleus)から構成されているので両者の総称として新線条体のことを Caudate-Putamen, CPu とも称する。
③A10の正式名称は腹側被蓋野です。作用は大脳辺縁系へ投射する中脳辺縁系路と前頭葉へ投射する中脳皮質路の二つがある。中脳辺縁系路は扁桃体の興奮(情動)により活性化され、中脳皮質路はストレスや不安等負の要因で活性化。
次にA11~A15までは間脳と言われる視床と視床下部のドパミン神経核の神経集団名A118~A15の五つの神経集団の正式な名称と主なドパミン作用を見ましょう。
④A11の正式名称は間脳後部です。作用は視床下部へ投射。レストレスレッグス症候群の関連部位
⑤A12の正式名称は弓状核です。作用は下垂体ホルモンの分泌を制御する。この系の抑制はプロラクチンの分泌を促進する(D2ブロッカーの高プロラクチン血症の原因部位)。
⑥A13⑦7⑧A14の正式名称は二つ合わせて不確帯です。 作用は視床下部へ投射
⑨A15の正式名称は嗅球です。
ドパミンの神経群として最も重要なのが、下図のA9:黒質緻密部とA10:腹側被蓋野の2つである。
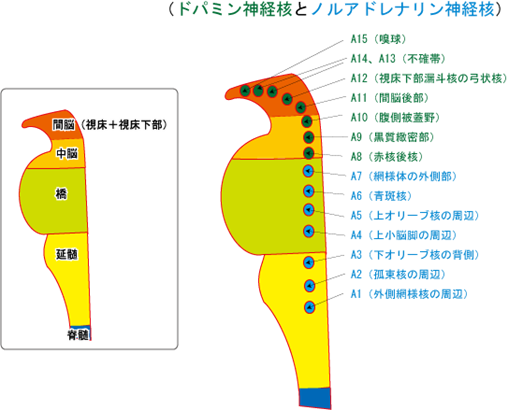
ドパミンはドパミンβ水酸化酵素の働きで、ノルアドレナリンにも合成される。ノルアドレナリンはCOMT(カテコール-O-メチル転移酵素)によってノルメタネフリンへ、MAO(モノアミン酸化酵素)によって、MHPG(3-メトキシ-4水酸化フェニールグリコール)へと分解され代謝される。
ドパミンの神経核
ドパミンの神経群として重要なのが、A9:黒質緻密部とA10:腹側被蓋野の2つである。
ドパミンの作用
A9:中脳黒質-線条体路
黒質-線条体路では、中脳の黒質に細胞体を置くドパミンニューロンが、線条体でGABAを伝達物質とする抑制性の中型有棘神経細胞を抑制的に制御しています。
この抑制制御がうまく行かない上に、アセチルコリン神経の活性化によりGABA神経の興奮→視床下核の興奮→視床の抑制→錐体外路障害を生じるのがパーキンソン病となります。
統合失調症では、主として前頭葉のNMDA受容体やドパミン受容体の異常によってドパミンの分泌が亢進し、ここ(基底核)のD2受容体を刺激してしまうため、視床が興奮して、ちょっとのことでもビクッとしてしまうような知覚過敏の状態を作り出してしまいます。
A10:中脳腹側被蓋野(中脳辺縁系路・中脳皮質路)
中脳腹側被蓋野から投射されるドパミンニューロンは大きく、腹側被蓋野-辺縁系路(側坐核・海馬・扁桃体等)と腹側被蓋野-皮質路(前頭前野)の2つに分けることができます。
中脳辺縁系路は正の強化因子、快情動、報酬系に関わる情動回路を動かすドパミンを分泌する経路、統合失調症では陽性症状の発現に関わっています。
さすってもらったら痛みが収まった→快情報として扁桃体が認識→記憶として海馬へ格納。この快の記憶が原因で扁桃体がA10を活性化→A10活性化によるドパミンが側坐核shellを興奮させて、視床内背側核→前頭前野→というYakovlevの情動回路を作動させて、もういちどさすって貰いたい!さすってもらえば痛みが収まる!といった快の情動行動を起こす方向へと進む。
中脳皮質路のほうは、不安やストレスにより活性化し、負の行動を起こす陰性症状の発現に関わっています。
ストレス刺激がA10経路を活性化すると、前頭前野のドパミンD1受容体を介して前頭前野の行う認知・思考・ワーキングメモリ等の機能を抑制してしまいます。
ドパミンの自己受容体
D2受容体には、D2L受容体とD2S受容体のサブタイプが存在している。
D2L受容体は、辺縁系では後シナプス、線条体では中型有棘細胞の後シナプスに多く発現している。D2S受容体は自己受容体として黒質、腹側被蓋野のドパミン神経終末(前シナプス)に発現している。
一方で、中脳皮質路のドパミン神経においては自己受容体が欠如しているため、ストレス刺激によって活性化してしまうとドパミンの分泌が制御できなくなるという。
脳波や目の動きなど生理的機能の研究からわかったことは統合失調症の患者さんは振り子を見てもらったときに、目の動きがぎこちなくなることや、特定の図形を見てもらったときに視点があまり動かず固定しがちであることなどが明らかになっていました。最近では、いろいろな刺激を与えた後に出現する脳波をコンピュータで分析し、事象関連電位という脳波上の微妙な変化を抽出することによって、刺激が脳の中でどのように情報処理されているかを調べる方法で、聴覚刺激を突然与えた時、約300 ミリ秒後に出現するP300という脳波の波形がよく調べられています。P300は聴覚刺激後脳内で複雑な情報処理を経てから出現するものですが、統合失調症の患者さんではP300の出現がよくなかったり、出現が遅れたりすることから、脳の情報処理過程の障害が推測されています。幻聴が多いのはP300の出現がよくなかったり、出現が遅れたりすることに関りがあるかもしれません。
統合失調症での認知機能の障害も調べられています。統合失調症の患者さんの認知機能のテストでは注意力・記憶力・言語能力などが低めで、抽象的な思考も苦手であることがわかります。更に行動の目標を設定し、柔軟かつ計画的に考え実行する力である実行機能も低下しています。 実際に、このような力が低下しているのは、統合失調症の患者さんの生活の中でしばしば観察され、社会の中で生活する上での障害になっています。
統合失調症での神経伝達の障害説には①統合失調症のドパミン仮説と②統合失調症のグルタミン酸仮説の二つがあります。
①統合失調症のドパミン仮説とは何でしょうか?統合失調症では治療薬の作用を手がかりにして研究が行われ、 統合失調症に対しては抗精神病薬とよばれる薬が有効であります。 この抗精神病薬が脳内でどのように働いているかを調べることは、動物を使っても研究できます。抗精神病薬は神経伝達物質であるドパミンの受容体2型とよばれる受容体を遮断します。つまり、ドパミンが受容体に働いて次の神経細胞に情報を伝えるのを、抗精神病薬は遮断しているのです。このことから、逆に統合失調症ではドパミンの機能が亢進しているのだろうと推測されました。これを統合失調症のドパミン仮説といいます。
ところで、ドパミンにはいくつかの経路があります。この経路のうち、統合失調症の病態に関連しているのは、中脳辺縁系あるいは中脳皮質系とよばれる経路です。 中脳辺縁系は腹側被蓋野とよばれる中脳の部位から大脳辺縁系に向かっています。 この経路は統合失調症の幻覚や妄想に関連していると考えられています。
一方、中脳皮質系は、同じ腹側被蓋野から前頭葉や側頭葉に向かっています。 統合失調症の陰性症状である感情の平板化、会話内容の乏しいこと、自発性の低下や社会からの引きこもりなどに関係しているのはこの経路ではないかといわれています。
黒質線条体経路は統合失調症の病態と直接の関係はないようです。ただし、この経路はパーキンソン病の病変部位です。従来からの抗精神病薬はドパミン経路のすべてに働いてしまうため、黒質線条体の遮断によってパーキンソン病様の症状である筋肉のこわばりやふるえなどの 錐体外路症状が副作用として現れやすかったのです。 覚せい剤やコカインを長期に服用すると、幻覚や妄想が現れることはよく知られています。覚せい剤やコカイン どちらの薬物も脳内のドパミンを増やし、ドパミンの機能を過剰にする作用を持っているので、先ほどのドパミン仮説にうまく合います。しかし、PETなどを用いて統合失調症の患者さんの脳内のドパミン受容体を測定しても、かならずしも受容体が増えているとは限らないようです。
②統合失調症のグルタミン酸仮説とは何でしょうか?脳内の神経伝達物質の機能障害という点では、グルタミン酸とよばれる神経伝達物質の機能障害も、統合失調症の原因として考えられています。 これは、フェンシクリジンとよばれる乱用薬を使用した人に、統合失調症と同じ症状が現れることから考えられました。
フェンシクリジンはグルタミン酸の受容体のうち、NMDAとよばれる受容体の機能を阻害するので、NMDA受容体を介したグルタミン酸機能の低下が統合失調症の原因ではないかというのがグルタミン酸仮説です。 ドパミン仮説が主に統合失調症の陽性症状を説明するのに対し、このグルタミン仮説は統合失調症の陽性症状と陰性症状の両方を説明できるのが特徴です。
ただし、この両者は相反する仮説ではなく、統合失調症の病態仮説についての2つの異なった切り口ととらえるべきでしょう。
統合失調症の成因(原因)についての仮説には①統合失調症の原因の神経発達障害仮説と②統合失調症の原因の神経変性仮説の二つがあります。
①統合失調症の原因の神経発達障害仮説とは何でしょうか?統合失調症の病的な状態を説明するものとして、ドパミン仮説やグルタミン仮説を説明してきました。 それでは、統合失調症はどのような仕組みで発症するのでしょうか。統合失調症の成り立ちについての仮説で有名なものに、神経発達障害仮説があります。
統合失調症では出生前後からのヒトの神経系の発達に何らかの障害があり、発症脆弱性といわれる病気のなりやすさが形成されると考えます。 このように脆弱性を持った子供が、思春期以降外界からのさまざまな心理的社会的なストレスを受けると統合失調症がはじめて発症するとされるのです。
この仮説を支持する所見として、統合失調症の患者さんは出産前後に産科的な障害を経験する率が高いことや、脳の海馬や内嗅皮質とよばれる部位などで神経細胞の発達が障害された痕跡のあることなどがあげられています。脳の海馬は「馬の脳」と呼ばれる「大脳辺縁系(だいのうへんえんけい)」に、「海馬(かいば)」という部分があります。 海馬は、タツノオトシゴのような形をしています。 日常的な出来事や、勉強して覚えた情報は、海馬の中で一度ファイルされて整理整頓され、その後、大脳皮質にためられていくといわれています。内嗅皮質は内嗅皮質ともいわれ海馬と共に記憶に深く関わる脳領域であり、双子のように類似した構造を持つ2つの領域、内側嗅内皮質と外側嗅内皮質から構成されます。 内側嗅内皮質は空間情報を処理する一方、外側嗅内皮質の細胞は匂いや物体の情報処理に関わります。内側嗅内皮質と外側嗅内皮質のどちらも、神経細胞が規則正しく整列した層構造から形成されており、層ごとに異なる配線を持ち、今述べたように異なる情報処理を行なっています。
また、この神経発達障害仮説によれば、統合失調症が思春期以降に発症しやすいことも説明できます。神経発達障害仮説では、患者さん自身の持っている脆弱性に、環境からのさまざまなストレスが加わり、はじめて発症するという、統合失調症の複雑な発症過程を説明できることが特徴です。
②統合失調症の原因の神経変性仮説とは何でしょうか?統合失調症でも進行すると認知機能の障害が見られます。統合失調症の多くの患者さんは治療に反応して症状は改善しますが、再発が多いのも事実です。再発の数が繰り返されると、病的な状態にある期間が長いほど、自発性の低下や社会からの引きこもりなどの後遺症に相当する症状が重くなっていきます。症状の重い患者さんほど、徐々に大脳が小さくなっていくようです。アルツハイマー認知症の患者さんのように目で見て明らかに萎縮していくというのではありませんが、アルツハイマー認知症と同じような病的な神経変性過程が統合失調症でも速さの違いはあってもゆっくりと進行しているようなのです。この仮説を統合失調症の神経変性仮説といい、最近注目されています。それは、早期に病気の治療を開始し、その後も再発を防ぐことができれば、このような神経の変性過程を停止させることができ、病気をそれ以上進行させないことにも繋がるからです。
延 髄 ( 自律神経、嘔吐、嚥下、唾液、呼吸の中枢部位 )
延髄(えんずい)は、脳幹のうちもっとも尾側の部分であって、吻側に橋、尾側に脊髄があります。後頭骨に開いた大後頭孔という穴を通る。背側には下髄帆を挟んで小脳があります。嘔吐、嚥下、唾液、呼吸および循環、消化の中枢を含み、生命維持に不可欠な機能を担っていて自律神経の中枢です。
循環中枢は交感神経、副交感神経の緊張度を調節することで心臓、血管の収縮を調節することにより血圧を制御しています。 血圧が急激に変化した場合には循環反射 circulatory reflex という機構が作動し、血圧の恒常性が維持されます。脳幹は、間脳、中脳、橋、延髄に区分される。
間脳は視床と視床下部からなる。視床は感覚神経を中継する場であり、視覚、聴覚などの情報を大脳皮質に伝える。視床下部は、自律神経機能の高位中枢である。
延髄には、心臓中枢、血管運動中枢、呼吸中枢、嚥下中枢など生命維持に重要な中枢がある。
脳幹に網目状に分布している脳幹網様体は、①筋の緊張や運動調節、②大脳にインパルスを送り、覚醒状態を保つ役割をしている。この上行性網様賦活系機能を抑制すると睡眠に陥る。小脳は、平衡機能、姿勢機能、随意運動などの調節を受けもっている。
ドパミンの神経核
ドパミンの神経群として重要なのが、A9:黒質緻密部とA10:腹側被蓋野の2つである。
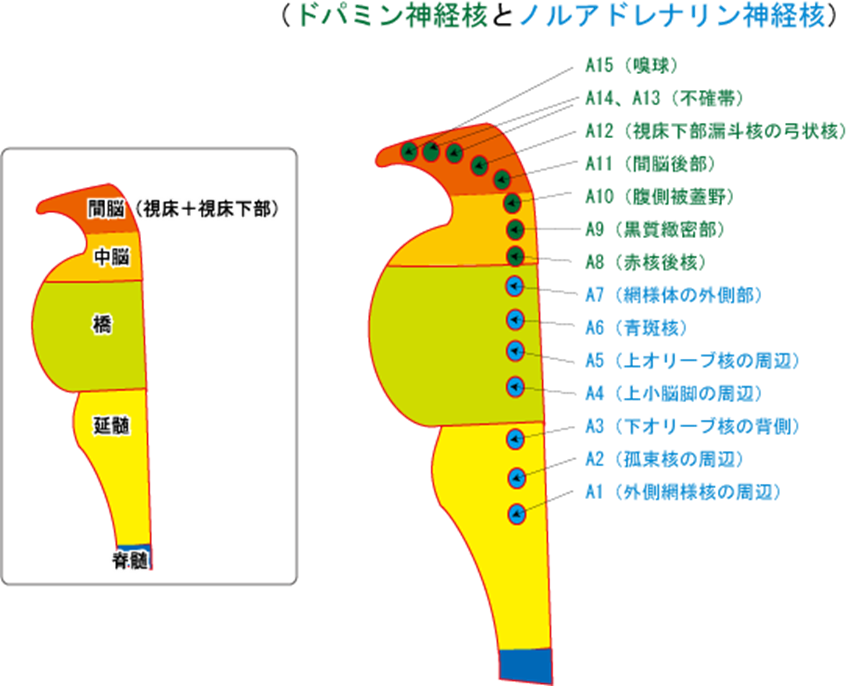
| 神経集団番号 | 神経集団名称 | 主な作用 |
| A8 | 赤核後核 | 辺縁系へ投射 |
| A9 | 黒質緻密部 | 新線条体へ投射。パーキンソン病の関連部位として錐体外路運動に関与 |
| A10 | 腹側被蓋野 | 辺縁系へ投射する中脳辺縁系路と前頭葉へ投射する中脳皮質路がある。前者は扁桃体の興奮(情動)により活性化され、後者はストレスや不安等負の要因で活性化。 |
| A11 | 間脳後部 | 視床下部へ投射。レストレスレッグス症候群の関連部位 |
| A12 | 弓状核 | 下垂体ホルモンの分泌に制御する。この系の抑制はプロラクチンの分泌を促進する(D2ブロッカーの高プロラクチン血症の原因部位)。 |
| A13、A14 | 不確帯 | 視床下部へ投射 |
| A15 | 嗅球 |
ドパミンの作用A9:黒質-線条体路
黒質-線条体路では、中脳の黒質に細胞体を置くドパミンニューロンが、線条体でGABAを伝達物質とする抑制性の中型有棘神経細胞を抑制的に制御しています。
この抑制制御がうまく行かない上に、アセチルコリン神経の活性化によりGABA神経の興奮→視床下核の興奮→視床の抑制→錐体外路障害を生じるのがパーキンソン病となります。
統合失調症では、主として前頭葉のNMDA受容体やドパミン受容体の異常によってドパミンの分泌が亢進し、ここ(基底核)のD2受容体を刺激してしまうため、視床が興奮して、ちょっとのことでもビクッとしてしまうような知覚過敏の状態を作り出してしまいます。
A10:腹側被蓋野(中脳辺縁系路・中脳皮質路)
中脳腹側被蓋野から投射されるドパミンニューロンは大きく、腹側被蓋野-辺縁系路(側坐核・海馬・扁桃体等)と腹側被蓋野-皮質路(前頭前野)の2つに分けることができます。
中脳辺縁系路は正の強化因子、快情動、報酬系に関わる情動回路を動かすドパミンを分泌する経路、統合失調症では陽性症状の発現に関わっています。
さすってもらったら痛みが収まった→快情報として扁桃体が認識→記憶として海馬へ格納。この快の記憶が原因で扁桃体がA10を活性化→A10活性化によるドパミンが側坐核shellを興奮させて、視床内背側核→前頭前野→というYakovlevの情動回路を作動させて、もういちどさすって貰いたい!さすってもらえば痛みが収まる!といった快の情動行動を起こす方向へと進む。
中脳皮質路のほうは、不安やストレスにより活性化し、負の行動を起こす陰性症状の発現に関わっています。
ストレス刺激がA10経路を活性化すると、前頭前野のドパミンD1受容体を介して前頭前野の行う認知・思考・ワーキングメモリ等の機能を抑制してしまいます。
ドパミンの自己受容体
D2受容体には、D2L受容体とD2S受容体のサブタイプが存在している。
D2L受容体は、辺縁系では後シナプス、線条体では中型有棘細胞の後シナプスに多く発現している。D2S受容体は自己受容体として黒質、腹側被蓋野のドパミン神経終末(前シナプス)に発現している。
一方で、中脳皮質路のドパミン神経においては自己受容体が欠如しているため、ストレス刺激によって活性化してしまうとドパミンの分泌が制御できなくなるという。カテコール核を持つアミン(カテコールアミン)で、中枢神経系の伝達物質、及び末梢のシグナル分子として働く。生体内のドパミンはチロシンから二段階の酵素反応によって合成され、小胞モノアミントランスポーターによって細胞内の小胞に取り込まれる。開口放出によって放出されたドパミンは放出部位から比較的離れた場所に存在する受容体に結合して標的細胞の生理機能を調節する。ドパミン受容体は全てGタンパク質共役型で、遅い信号伝達もしくは神経細胞機能の修飾を担う。中脳から大脳に投射するドパミン神経が中枢のドパミン神経系の大部分を占め、運動機能、認知機能などの中枢機能の調節に関与する。また、ドパミン神経系は精神疾患の病態生理に対する関与が示唆されており、抗精神病薬等の治療薬や依存性薬物の標的となる。
