ミトコンドリアの機能障害によって人体のエネルギー通貨であるATPの産生低下ために癌が生じるという理論は間違いであることを証明します。まずミトコンドリアでの活性酸素の発生と除去について詳しく述べます。
生きるために絶対に必要な酸素の消費に必然的にミトコンドリアで生み出される活性酸素によって仮にDNAが傷害された時にDNAの修復のためにDNAの修復系が生まれたのです。DNAの修復系によってDNAの修復が不可能であるときにはアポトーシス(細胞自殺)という二段構えの最終手段を用いて傷ついた細胞自身にミトコンドリアが責任を取ってミトコンドリア自らが強制する自殺なのです。
活性酸素は恐れる必要はありません。活性酸素が癌の原因といわれますが癌を生み出すのは活性酸素ではなく細胞のゲノムに感染したヘルペスウイルスが増殖遺伝子を変異させたために増殖過剰遺伝子に変えたために異常に増殖過剰細胞になったのです。増殖過剰細胞に変えたために癌といわれる病気が生まれるのです。癌細胞などは作られてしまった「癌即死ぬ」という強烈なイメージが恐ろしいだけ実際は癌は何も怖くはないのです。一番怖いのは現代に残されたすべての病気を作るヘルペスウイルスなのです。もっと怖いのは嘘で患者をだまして治療と称する免疫を抑制して症状だけ軽減するだけでヘルペスウイルスを増やし続けて病気を治すという責任を放棄してお金を稼ぎ続ける医薬業界なのです。資本主義体制が持つ矛盾そのものです。残念です。
人体のエネルギー通貨であるATPを作り出し、かつアポトーシスを生み出すミトコンドリアとは、またアポトーシスとは何でしょうか?
アポトーシスapoptosisは、apo(off、離れる)とptosis(falling、落ちる)を合成した「遺伝子に組み込まれた細胞死」の術語です。アポトーシスは、生体の中で不要となった細胞を除去する生体制御機構であり、突然変異や傷害を受けて異常となった細胞を排除する生体防御の意義も兼ね備えている。すなわち、正常・異常細胞の自己消去機能による”生の更新”がアポトーシスの本質といえる。
アポトーシスはネクローシスnecrosisと異なる細胞死として形態学的観察から提起されたため、その検出も当初は核の形態学的変化(核周辺部へのクロマチンの凝縮margination)に基づいて行われた。
核周辺部へのクロマチンの凝縮marginationとは何でしょうか?クロマチン凝集とは、細胞の核内のクロマチンが凝縮する現象を指します。クロマチンは、DNAとタンパク質の複合体で、細胞核内のDNAを効率的に詰め込む役割を担っています。クロマチン凝集は、細胞の分裂やアポトーシス、放射線照射など、さまざまな場面で起こります。クロマチン凝集の役割は、細胞分裂の際に、クロマチンが凝縮してよく見られるようにXの形をしたゲノムDNAの構造となります。アポトーシス(細胞死)の過程で、核内のクロマチンが凝縮し、核の断片化や細胞の分断化などの劇的な変化が起こります。又放射線照射によってDNAが損傷を受けるのを防ぐために、クロマチンを凝縮させています。真核生物では、次世代に伝える遺伝子情報を安全に維持するために、クロマチンを凝縮させています。
クロマチン凝集の制御は重要で、クロマチンの凝集状態の違いによってゲノムDNAからの読み取りができるかどうかが決まります。エンドヌクレアーゼ活性によるヌクレオソーム(約180 塩基対)単位のDNA断片化がアポトーシスを特徴づける。
ヌクレオソームとは、真核生物の細胞核内でDNAがとる基本構造単位で、核たんぱく質の構造体です。ヌクレオソームの構造は①中央に4種類のヒストンタンパク質(H2A、H2B、H3、H4)が2分子ずつ集まったヒストン8量体がある②その周囲にDNA二重らせんが約1.5回ほど巻き付いている。
ヌクレオソームの役割は①DNAを保護する②DNAの情報を利用できるようにする③遺伝子の活動を部分的に変更する。
クロマチンとはヌクレオソームが連なった構造をクロマチンと呼び、DNAの配列と構造を保持しながら効率的に核に詰め込む役割があります。クロマチン構造は動的に変化することで、遺伝子の発現のスイッチがオンになったりオフになったりします。アポトーシスの特徴はネクローシスと違ってこのようなDNAの断片化による染色体の処理が行われるのです。
アポトーシスの過程において生じるエネルギー依存性の生化学的・形態学的変化の多くは、カスパーゼcaspase類による特定の蛋白質の限定分解による。切断・活性化されたcaspase 3は、poly (ADP-ribose) polymerase (PARP)、cytokeratin 18(CK18)やactinといった細胞内蛋白質を限定分解する中心的役割を担う。サイトケラチン18(CK-18)は肝細胞などの細胞骨格を構成する蛋白の一つで、肝細胞のアポトーシス. により断片化されます。Cytokeratin 18(別名:KRT18、CYK18、PIG46、K18)は、中間径フィラメント遺伝子ファミリーで最もよくみられるメンバーの一つです。
カスパーゼとは何でしょうか?アポトーシスの主要な実行過程はDNA断片化とカスパーゼによる蛋白質の限定分解カスケードであり、アポトーシス制御蛋白質であるp53蛋白、Rb蛋白、bcl-2蛋白、bcl-xL蛋白、bax蛋白、c-myc蛋白、cyclin類などが関わっています。
アポトーシスが起こる仕組みとは。
カスパーゼの活性化を引き起こすのはミトコンドリアです。どのようにしてミトコンドリアでカスパーゼの活性化が引き起こされアポトーシスが起こるのかの仕組みを詳しく解説しましょう。
人体の細胞は増殖能から区別できる3種類の細胞群に分けられます。この細胞の増殖能が癌の生りやすさを決めているのです。①常に増殖をくり返しているrenewing cell(腸上皮、表皮、造血細胞、精母細胞、癌細胞など)、②ふだんは増殖しないが必要に応じて増殖できるstabilized cell(線維芽細胞、血管内皮細胞、尿細管上皮細胞、肝細胞など)、そして③増殖能のないstable cell(神経細胞、卵細胞、心筋細胞など)の3種の細胞群に分けられます。
因みに増殖能を持っている①と②の細胞群は分裂増殖するときにヘルペスが細胞のゲノムに自分のゲノムを組み込み増殖遺伝子を変異させて発癌させる可能性を増やすことができるのです。ところが③の増殖能のないstable cellである三つの細胞である神経細胞、卵細胞、心筋細胞は増殖する必要がないのでヘルペスウイルスはこれらの細胞のゲノムに入り込めないので発癌が不可能なので癌は生じません。興味あることには精母細胞には癌が起こりえますが、卵細胞には癌が起こらないことです。なぜなら、renewing cellでは増殖速度に応じたアポトーシスが生理的に(恒常的に)生じているのでヘルペスが細胞のゲノムに感染しやすくなり増殖遺伝子が変異しやすくなり発がんの可能性が高くなるのに対して、stable cellにおけるアポトーシスすることは絶対ないうえに発癌も起こらないのです。
胚中心細胞とは胚中心細胞とは、リンパ節や脾臓などの免疫組織において、免疫応答中に濾胞(ろほう)内に作られる反応場である胚中心で増殖するB細胞です。
胚中心では、増殖するB細胞が自己の抗体遺伝子に変異を入れることで、抗原に対する親和性を向上させます。この反応を胚中心反応と呼び、抗体が抗原と結合しやすくなる性能を高め、生体防御の役割を果たします。
胚中心は、細菌やウイルスなどの感染によって形成される一過性の構造です。Bリンパ球が集まる場所を濾胞と呼び、胚中心は濾胞の中の、Tリンパ球の領域に近接した部分に形成されます。
卵細胞(らんさいぼう)とは、雌の生殖細胞で、生物における雌性の配偶子です。脊椎動物では「卵」や「卵子」とも呼ばれます。卵細胞の特徴は①一般的に球形で、卵黄を含んでいる②精子と比べて大型で運動性が低い③種によって卵黄の量や分布が異なる④極性を示し、動物極と植物極が区別される⑤卵巣内で形成され、成熟のときには卵母細胞の減数分裂によって生じる⑥染色体は半減している。
女性が一生で排卵する卵子の数は400~500個です。思春期から閉経までの40年間で月一回の排卵のチャンスがありますから40×12で500となります。
女の赤ちゃんは出生時には、卵子の元である卵母細胞を100~200万個を持っていますが、思春期頃には30万個まで減少します。
植物の卵細胞は、子房にある胚珠と呼ばれる組織の中でつくられます。花粉症のシーズンに飛び交う花粉とは、植物のおしべにある葯(やく)と呼ばれる袋の中で作られる粉末状の物質で、植物の繁殖に重要な役割を担っています。おしべが作る花粉には、植物が子孫を残すためのDNAが詰まっており、風や昆虫によって運ばれてめしべに受粉することで種が作られます。花粉の形態は植物の科や属によって異なり、発芽口の形や数、粒数、気嚢数などで分類されます。受粉すると精細胞が入った花粉管が胚珠に向かって伸びていき、精細胞のうち1つだけが卵細胞に到達して受精卵ができます。

ミトコンドリアは、細胞内に存在する細胞内小器官で、細胞のエネルギーとなるATP(アデノシン三リン酸)を生成する役割を担っています。また、脂質代謝や病原体を殺傷してしまう炎症反応、人体に迷惑をかける異常な細胞をアポトーシスといわれる遺伝子に刻印されているので計画された細胞死と呼ばれるシグナルを送り実行するなどの、さまざまな生命を守るために必須な機能も果たしているのです。
ミトコンドリアは、食事から得られるブドウ糖と呼吸から得られる酸素を使って最も多くのATPを生成します。この仕組みを「好気呼吸」といいます。「酸素を好む呼吸」という意味です。
ミトコンドリアは、人類が誕生した約20億年前に真核細胞の祖先が酸素を用いてエネルギーを作る細菌を取り込んだことで誕生したのです。そのため、ミトコンドリアには細菌由来のDNA(ミトコンドリア、英語で単数形で mitochondrion、複数形で mitochondria、略して、mtDNA)が存在します。
ミトコンドリアの機能が低下すると、人体のエネルギー通貨と言われるATPの生成量が減り、同じ量のエネルギーを作るためにより多くのエネルギー源を摂取する必要があります。また、ミトコンドリアの働きが低下すると、細胞の活動が低下し、エネルギー不足でさまざまな症状が現れます。たとえば、脳の神経細胞であれば見たり聞いたりすることが障害され、心臓の細胞であれば血液を全身に送ることが困難になります。人体にはすべての細胞にミトコンドリアが存在しますが、特に腸と腎臓に多く存在しています。
ミトコンドリアの機能が低下するのは、、ミトコンドリアの機能が低下する原因には、①ストレス②運動不足③herpesによる遺伝子の変異、④加齢、⑤過労、⑥睡眠不足、⑥喫煙、⑦飲酒、⑧活性酸素、⑨低血糖、⑩質的栄養不足(ビタミン、ミネラル不足)、⑪ヘルペスによる慢性炎症、⑫有害金属の蓄積、⑬カビ毒(腸カンジダなど)、⑭腸内環境の乱れなどです。つまり不健康な生活のすべてがミトコンドリアの機能が低下をもたらします。
ミトコンドリアの機能の低下が癌の原因という人たちがいますが不健康な生活のすべてが免疫を下げてしまうとヘルペスが増えて感染細胞が多くなるにつれてゲノムの増殖遺伝子を変異させてしまう確率が増えて発がんが始まるから悪性腫瘍の原因はやはり漠然とした不健康な生活が何となく癌を生むのではなく不健康な生活に潜む免疫の低下が癌ウイルスであるヘルペスウイルスを増やし細胞の増殖遺伝子を変異させてしまうからです。
人体には40兆の細胞があり、1個の細胞の23500個の中には増殖関連遺伝子がその5%の約800個あります。この正常な増殖関連遺伝子の800個の内、ヘルペスウイルスが変異させてしまう増殖関連遺伝子が多ければ多いほど悪性度の高い癌の進行度も高度となる増殖過剰細胞(癌細胞)が次々と増えていく可能性も増えて次々と過剰増殖細胞(癌細胞)が蓄積していき、と同時に増えたヘルペスはまだ感染細胞や、すでに癌細胞になってしまった細胞でも居座り続けて、細胞が取り込んだ酸素や5大栄養素を奪い取り無限大にヘルペスビリオンを生み出すために癌細胞も正常細胞も飢餓状態になり痩せ衰えてしまい最後は多くの正常な細胞も栄養不足になります。人体のホメオスターシスを維持できなくなり細胞栄養失調症になっても食べれば食べただけ栄養素やエネルギーを増殖の勢いの強いヘルペスビリオンにかすめ取られ続けて最後の最後は多くの細胞の餓死となり多細胞で成り立っている人体にも死が訪れるのです。
癌の人が死ぬのは「癌ではなく」増えすぎたヘルペスによって栄養を奪われて食物が満ち溢れている現代ではめったに見られない極端なマラスムスとクワシオルコルで飢え死に同然の死に方である悪液質でとどめを刺されてしまうのです。つまり癌による「餓死」は普通の「餓死」ではないのです。食べ物も飲料水も無限にあるのですが細胞の正常な活動に必要な飲食物を摂取しても増え続けるヘルペスビリオンが横取りしてしまうので正常な細胞を構築できなくて細胞の物質代謝やエネルギー代謝も阻害され続きます。細胞を顕微鏡で見ると顔つきの悪い出来の悪い細胞ばかりとなり「このままでは癌死するよと脅されて治すことのできない後遺症で悩むだけの手術を勧められて早死にすることにもなります。躊躇しているとセカンドオピニオンを紹介されても同じ標準医療を行うだけですから前医と同じ治療法を勧められるだけですから「セカンドオピニオン」も「同じ意見」となりますから無駄なことになるのです。
悪液質とは何でしょうか?癌の原因であるヘルペスビリオンが無限に増や続けるために体内で人の体の機能と構造のほとんどすべてに関わるたんぱく質が合成できず、脂肪量の減少の有無に関わらず逆に筋肉内の蛋白質もherpesの増殖のためにエネルギーや構造蛋白としても利用されるために破壊される複合的代謝異常の症候群で、栄養不良状態が生じ衰弱した状態」と定義されます。悪液質の主な症状は①体重減少、②骨格筋量減少、③食欲不振、④ 疲労やだるさ。herpesが人体に必要なエネルギーや栄養素を略奪されて生まれる悪液質は、がん患者に多くみられる合併症の1つで、特に進行した消化器がんや肺がんで高頻度に発症するのは当然です。
悪液質は「前悪液質」「悪液質」「難治性悪液質」の3つのステージに分類されますが、難治性悪液質まで進行すると効果的な治療方法がなく、予測生存期間は3か月未満です。悪液質(Cachexia)とは「ヘルペスの莫大な増殖のために体内でたんぱく質が合成できず、逆に筋肉内のたんぱく質までもヘルペスに奪われて破壊されることで栄養不良状態が生じ衰弱した状態」と定義されてきました。
がんは特別な病気ではないのですが、「通常の栄養サポートでは完全に回復することができず、進行性の機能障害に至る、脂肪量減少の有無にかかわらず、骨格筋量の持続的な減少を特徴とする生き続けるヘルペスウイルスを増やし続けるために癌死するまであらゆる栄養素をヘルペスウイルスに奪われる症候群」と定義してもよいのです。
クワシオルコルとマラスムスとは何でしょうか?栄養障害を示す言葉です。栄養とは、生物が外界から食物を取り入れて、成長や活動、生命維持に役立てる一連の活動です。栄養の源となる物質を栄養素といい、栄養素をバランスよく摂取することが健康を維持するために大切です。栄養素には、①炭水化物、②脂質、③タンパク質、④ミネラル、⑤ビタミン、⑥食物繊維、 ⑦水の7つです。食物繊維も水も栄養素の一つなのですよ。
五大栄養素とは、三大栄養素といわれる「炭水化物」「脂質」「たんぱく質」に「ミネラル(無機物)」「ビタミン」を加えたものです。各栄養素にはそれぞれ異なる働きがあり、その働きによって以下の3つのグループに分けられます。
栄養素の働きによる分類。
①エネルギーのもととなるのは、炭水化物、脂質、たんぱく質の3つです。
②体を作るのは、たんぱく質、ミネラルの2つです。
③体の調子を整えるのは ビタミン、ミネラルの2つです。
①の体に必要なエネルギーのもととなる主な栄養素は、三大栄養素である「炭水化物」と「脂質」「たんぱく質」です。臓器の基本的な働きを助け、脳と体を動かす力の源になることから、3つの栄養素は「エネルギー産生栄養素」とも呼ばれます。炭水化物は糖質と食物繊維に分けられ、糖質は日常的な活動に使われるエネルギー源のひとつです。脂質は体に蓄積されて効率良くエネルギーを生み出します。たんぱく質は、糖質や脂質が体内に不足している場合に分解されてエネルギー源になります。
②の体を作る働きをする栄養素は、「たんぱく質」と「ミネラル」です。たんぱく質は、臓器、皮膚、筋肉、血液、髪の毛など、体を構成するあらゆる成分の材料として使われます。
一方、ミネラルは、カルシウム、ナトリウム、カリウム、マグネシウム、亜鉛、鉄、銅、マンガン、リンなどの総称で、骨や歯を作る、神経伝達物質の合成を行う、身体機能の働きを助けるといった役割があります。
③体の調子を整える栄養素は、「ビタミン」と「ミネラル」です。体を動かすエネルギー源にはなりませんが、体の機能を調節して調子を整えます。
このように、五大栄養素には生命を維持し、健康的な生活を送るためのさまざまな役割があります。食事の際は五大栄養素をバランス良く取り入れるようにしましょう。
体内で合成できない栄養素は必須栄養素と呼ばれ、ビタミンやミネラル、一部の必須アミノ酸、一部の必須脂肪酸などです。
低栄養状態が必要とする栄養の摂取量を摂取できていない結果、身体的活動レベルが低下してしまい、血清アルブミン濃度が3.5g/dlになった状態を指します。栄養不良のことです。高齢者は嚥下機能・消化吸収機能が低下してくるために、なりやすくなります。
低栄養状態になると、活動性が低下、持病の悪化など、健康状態にも影響が出てきます。大きく分けるとマラスムスとクワシオルコルの二つがあります。
- マラスムスはガン末期患者のように、長期間、栄養補給がされずに、骨と皮だけにやせ細ってしまった状態のことです。癌を作り続けるヘルペスウイルスが増えれば増えるほど膨大な数になっていけばなっていくほど摂取されたすべての栄養素をヘルペスウイルスのビリオンにすべて奪われ続け最後は悪液質で死ぬまで長期間、低栄養状態が続き人体の細胞に補充されずに、骨と皮だけにやせ細ってしまいます。マラスムスは「エネルギーの欠乏」が主体となって起こる栄養障害です。勿論、同時に、たんぱく質摂取量も減少しています。特徴として、著明な体重減少などが見られます。
- クワシオコルはたんぱく質が摂取されず、浮腫が起きている状態のことです。開発途上国で多く見られ、腹水が溜まり、下腹部だけ膨らんでしまっていることが多いようですジブチでは通常、8 人に 1 人の子どもが 5 歳の誕生日を迎える前に命を落とし、10 人に 1 人の子どもが 1 歳になる前に死んでしまいます。
「たんぱく質の欠乏」が主体となって起こる栄養障害です。しかしエネルギー摂取量は比較的保たれています。特徴として、低栄養性の脂肪肝や低アルブミン血症、浮腫などが見られます。
クワシオコルで低栄養性の脂肪肝が起こるのは通常、肝臓の脂質は、リポたんぱく質という船に乗り末梢組織へ運搬されます。しかし、たんぱく質欠乏状態では、リポたんぱく質の形成が上手くできず、肝臓に脂質が蓄積し“脂肪肝”が起こります。
浮腫が起こるには、たんぱく質には、血管内に水を引きつけておく力(膠質浸透圧)があります。たんぱく質が十分あれば、血管内に水を保持しておくことができます。しかし、クワシオルコルのようにたんぱく質が欠乏してしまうと、
水を血管内に引っ張る力も弱くなってしまいます。これを膠質浸透圧の低下といいます。
その結果、血管外に水が漏れ出し“浮腫”が起こります。
がんの痛みはなぜ起こるのでしょうか?がんの痛みは、がんが進行し末期を迎えるほど激しくなることが多いですが、必ずしもそのように起こるわけではありません。がんの痛みは、癌の原因であるヘルペスが全身に分布しているポリモーダル自由神経終末の侵害(疼痛)受容器をもっている神経細胞に感染すると痛みがみられます。いずれにしろ痛覚神経にヘルペスが感染しなければ痛みは出ません。癌の原因であるヘルペスが多くなればなるほど痛みが出やすくかつ高度になります。初期がんではヘルペスが痛みの原因となり3分の1異常の患者が、末期がんでは80%以上の3分の2以上の人が痛みを感じます。
がんの痛みの発生時期は早い時期から出始めて末期に近づくほど痛みが激しくなることが多くなっていきます。癌が治るわけでもない三大標準医療をやればやるほど免疫が落ちすぎて癌の原因であるヘルペスが増えすぎて痛覚神経細胞にも冷えすぎて感染してしまうのです。ヘルペスが全身のポリモーダル自由神経終末の侵害(疼痛)神経細胞の受容器に感染するときに起こる神経因性疼痛が様々な臓器に見られます。主な痛みの種類は内臓痛では締め付けられるような鈍い痛み、骨転移の場合は体を動かすときに鋭い痛み、時に焼けるような、もしくは電気が走るような痛みがあちこちの臓器におこります。神経因性疼痛とは、ヘルペスによって痛覚神経が障害されることで生じる痛みですが癌の塊による圧迫痛といわれますが癌の塊は人体の細胞の塊に過ぎないので絶対に圧迫痛などが起こるわけはありません。ましてや灼熱痛やピリピリ感、ズキズキする痛み、鋭い痛みなどは痛覚神経にヘルペスが感染しているからです。がんの痛みは、焼けるような、うずくような持続する痛みや、ビーンと走るような、びりびりするような電撃痛などが特徴です。また、痛みがあるところに脱力(自由に動かせない)が伴うこともあります。
がんの痛みは難治性であることが多く、非オピオイドやオピオイドに加えて鎮痛補助薬(抗痙攣薬、抗うつ薬など)が必要となります。痛みが比較的軽度の場合は、アセトアミノフェンやNSAIDsなどの非オピオイドが使用され、中等度から高度の場合には、麻薬系鎮痛薬であるオピオイドが使用されます。
また、末期がんの患者は、痛みとは異なる苦痛として、耐えられないほどのだるさ(倦怠感)や腹部の膨満感、吐き気、嘔吐、息苦しさなどにも苦しむことが多くあります。
神経障害性疼痛は、ヘルペスウイルスによる帯状疱疹が治った後の傷ついた疼痛神経の長引く痛みや、糖尿病で免疫が落ちた後の合併症に伴う痛み、坐骨神経痛、頚椎症に伴う神経障害疼痛など、さまざまなヘルペス性の痛みが起こります。癌の場合の様に傷や炎症などが見えないにもかかわらず痛みがある場合、ヘルペスがポリモーダル自由神経終末の侵害(疼痛)神経の受容器に感染して炎症が原因となっているのです。癌細胞は本来自己の細胞が増殖しすぎたに過ぎないのにもかかわらず炎症が起こるのは正に癌細胞にはherpesウイルスがうようよと蔓延しているのでCRPや血沈やSAAなどの炎症所見が高くなるのです。「癌」とは簡単に言うとヘルペス感染症であり、難しく言うとherpesが細胞の核のゲノムにまで感染して癌関連遺伝子(増殖遺伝子)を変異させてしまっただけの細胞に過ぎないのです。
最後に皆さんに質問を出します。実は癌で死ぬのはヘルペスが癌関連遺伝子(増殖遺伝子)を変異させすぎてしまってヘルペスに栄養を略奪されて「極度の栄養不良」という曖昧な原因で死ぬのではなく本当に正しい答えは別にあるのです。この文章の最後に答えを書き記しましからそれまで、皆さんに考える時間を少しばかり差し上げますから答えを見る前にしっかり考えてください。痛みで死ぬこともないのは当然です。ヒントになる文をいくつか書き記します。
癌で死ぬ直前の症状には、次のようなものがあります。
- 呼吸の変化:呼吸が不規則になる、息苦しさを訴える、死前喘鳴(咽頭や喉頭部に分泌物が貯留して呼吸の際にゴロゴロと音がする)など。死前喘鳴とは死前喘鳴(しぜんぜんめい)とは、死亡直前(死亡数時間前から数日前)に生じる、呼吸に伴う不快な音のことです。Herpesが気道内に感染して気道内分泌物の貯留によって、咽頭や喉頭部の分泌物が振動して「ゼーゼー」「ゴロゴロ」という呼吸音になります。死前喘鳴は、終末期のがん患者では40~70%に生じる。死前喘鳴の原因は死ぬ直前のストレスと免疫を抑える間違ったがん医療のために咽頭や喉頭部の粘膜細胞に感染したヘルペスウイルスと、気道にも感染しているヘルペスと自然免疫による炎症のために蓄積した分泌物によって起こるのです。更に死期が迫り意識レベルが低下すると嚥下反射が抑制されるため、唾液分泌物が咽頭部に分泌物が蓄積するのです。嚥下反射とは、食べ物を食道に送り込むために起こる反射運動です。口腔内が密封された瞬間に起こり、喉の喉頭蓋が下がって気管への入り口をふさぎ、食塊が気管に入らないようにします。嚥下反射が弱くなると、飲み込みにくくなったり、誤嚥を起こしやすくなったりするなどのリスクにつながります。嚥下反射の惹起遅延とは、のどの感覚が鈍くなっているために、食べ物がのどに来た感覚が分かりにくく、嚥下運動がおこるまでに時間がかかることを指します。
死前喘鳴は、数時間ないし数日間で死に至る可能性があることを示す徴候です。患者さんにとっては苦痛ではないようです。
- 意識・認知の変化:寝言が増える、辻褄の合わない発言や幻覚・妄想がみられる、寝ている時間がだんだん長くなる、呼びかけても起きない昏睡状態へと至るなど
皮膚の変化:足が浮腫む、手足が冷たくなる(末梢冷感)、足の裏がまだらの紫色になる(チアノーゼ)、顔色が白くなる、口元がだらんとしてほうれい線(鼻唇溝)などが緩む。
- 目の色:栄養が摂れなくなると目の周りなどさまざまな箇所に影響が及び、目の色が濁りはじめる。ほうれい線とは、鼻の両脇から口角外側に伸びるシワや線で、医学用語では「鼻唇溝(びしんこう)」や「ナゾ・ラビアル・フォールド」と呼ばれます。加齢とともに目立ち始め、ほうれい線が濃くなると年齢を感じやすく、お化粧でも隠しづらくなります。
ほうれい線ができる原因としては、肌の張りの減少やホホのたるみなどが挙げられます。また、乾燥によって肌のバリア機能が低下し、外の刺激から受けるダメージが大きくなると、ほうれい線が深くなりやすいのです。
これらの兆候は医学的には早期・晩期死亡前兆候と呼ばれ、見た目や触ってわかる変化です。がんの部位や組織が違っても、終末期になるほど顕在化するという特徴があります。
亡くなる約1ヶ月前には、食欲不振や倦怠感、呼吸困難感などの症状が出現し、これらの症状は日を追うごとに増強していく傾向があります。 多くの場合、がんが進行しても患者様の全身状態はしばらくの間保たれています。 しかし、死亡が近づく約1ヶ月の間に、その全身状態が急速に低下するのが特徴的です。
どんな症状が出るかは病名や体の状態によって十人十色ですが、よく起こる症状としては、①倦怠感、②食欲不振、③痛み、④便秘、⑤不眠、⑥呼吸困難、⑦悪心嘔吐、⑧せん妄などがあります。悪心嘔吐とはストレスなどの精神的な影響により起こる吐き気には、自律神経が大きく関係しており、脳が精神的ストレスを感じると交感神経が優位になり、その状況を回避し、迅速に体を動かせるように、延髄にある嘔吐中枢が刺激されます。 それにより、嘔吐反射や絞扼反射が起こります。絞扼反射(異常絞扼反射とも呼ばれる)とは、吐き気をもよおす反射です。口の中(舌、軟口蓋、咽頭部、喉頭部など)の刺激によって誘発される、吐き気をもよおす反射です。有害な物質の侵入を阻止するための防御反応として機能します。悪心と嘔吐の違いは
下の表に各疾患でどんな症状がどれくらいの頻度で出現するか記載しています。例えば、がんでの倦怠感の出現頻度は23〜100%と大きな幅がありますが、これは文献によって報告内容が異なるからで、それくらい症状が出現するかどうかはわからない、ということになります。
ただし、きちんと薬を調整したり、環境を整えれば、ほとんどの症状は緩和できますので、「死ぬ時には苦しい思いをしなければならない」と怖がる必要はありませんどころか、「癌死」は癌で死ぬのではなく、ヘルペスウイルスの過剰増殖と患者からエネルギーと栄養素のすべて奪い取られてしまうので死ぬのです。決して癌細胞が過剰増殖したためでもなくと癌患者からエネルギーと栄養素のすべてを癌細胞に奪い取られてしまうので死ぬわけではないのです。癌の原因はヘルペスウイルスの過剰増殖なので原因がわかっているので癌は治せる病気なので何も怖がる必要がないのです。皆さんに「癌はヘルペスが原因であるので癌になりたくなければヘルペスを増やさないように心にストレスをかけて免疫を落とすなと伝えたいのです。他人の喜びを自分の喜びにしましょう。極めて簡単なことなのですよ。


緩和ケアは、歴史的にはがん患者に焦点をあてて発展してきました。本来、緩和ケアは疾患、病期、場所を問わず、すべての患者に必要とされるものです。近年、心不全、呼吸不全、神経難病などの非がん患者への緩和ケアの必要が認識されるようになり、日本でもその取り組みがはじまっていますが難病のほとんどがヘルペスが原因となっているので「ヘルペスウイルスを増やさないように免疫を落とすな」という教育が一番大切なのです。
がん患者の特徴はがん患者は、亡くなる2~3ヵ月前まで日常生活を大きな支障なく送ることが多いです。しかし、亡くなる約1ヵ月前になると、食欲不振、倦怠感、呼吸困難感などの症状が出現し次第に増強します。がんは進行しても全身状態は保たれますが、死亡前の約1ヵ月で急速に全身状態が低下することが特徴です。がんの部位や組織が違っても、症状や臨床経過において、一定の共通性や規則性が認められ、終末期になるほど顕在化するという特徴をもちます。
身体症状の出現からの癌死までの生存期間の痛みと緩和医療については癌死までの生存期間が1ヵ月以上の場合、痛みの出現頻度がもっとも高く、生存期間が1ヵ月頃から倦怠感、食欲不振、便秘、不眠などが増加する傾向があります。生存期間が2週間頃よりせん妄が増加しはじめ、死亡数日前より不穏や死前喘鳴(咽頭や喉頭部に唾液や気道からの分泌物が貯留し、呼吸の際にゴロゴロと音がすること)がみられるようになります。死が近づくほど、一人の患者により多くの症状が現れ、またそれぞれの症状はより緩和困難となってきます。このように、がんの終末期に現れる症状の出現頻度と出現時期には一定の傾向がみられ、最後の1ヵ月はとくに重点的な症状の緩和が必要となります。
因みにLynnは、終末期の疾患の軌跡を、①がんなどの進行性疾患のモデル、②心不全、COPDなどの臓器不全モデル、③認知症・老衰モデル、の3つに分類しました。癌死は②のCOPDと同じく肺機能不全による死と述べていますが肺癌ではほとんどが肺機能不全ですから癌死の原因も癌の種類によって悪液質ではない死に方があるのです。
最後になりましたが癌死の真実の直接の原因に対する答えは以下に書きます。
「癌死」の最終的な直接的な原因は肺癌でも肺の自律神経の交感神経節に感染したヘルペスが大量に神経細胞体に感染しまくって神経筋シナプス伝達が不可能になって肺が呼吸できなくなり、酸素と二酸化炭素の入れ替えが不可能になり呼吸不全になったためですと言うべきです。酸素と二酸化炭素を入れ替える機能を持っている肺の1型肺胞細胞のすべてが肺癌細胞になっているわけでないからです。
Herpesによる神経障害性疼痛は、通常の痛み止めの効果が期待できず、難治性の痛みになります。治療には、薬物療法や理学療法、外科治療、麻薬の投与などが行われます。
神経障害性疼痛には、発作性神経痛と持続性神経痛の2種類があります。発作性神経痛は三叉神経節に隠れ潜んでいる増えたヘルペスが免疫細胞に見つかって生じた炎症による三叉神経痛などが代表例で、癌末期によくみられるherpesウイルスによる持続性神経痛は24時間絶え間なく痛みが持続するので麻薬を使わざるを得なくなるのです。がんの原因はヘルペスであるので癌に関わる症状はすべて痛みも含めてヘルペスと関連している症状であるのです。
Herpesが起こすがんの痛みを和らげる医療として「緩和ケア」があり、痛みや苦しみを和らげることを優先して行われるので免疫を確実に抑制してしまう麻薬を使います。
また、がんの痛みだけでなく、手術、放射線治療、化学療法などの治療に伴う痛みや入院生活中におこる筋肉痛や褥瘡などもすべてヘルペスによる痛みなのです。
ミトコンドリアについての知識をさらに深めましょう。まず最初に「ワーブルグ効果」とは何かについてから始めます。次に「ワーブルグ効果」と「癌」についてのいくつかの論文を勉強しながら正常細胞を癌細胞に変異させるヘルペスウイルスを介在させると「癌」についての矛盾が解明可能となるので「ワーブルグ効果」の誤りを正しましょう。
「ワーブルグ効果」のすべてが誤っているにではなく「がん細胞が正常細胞とは異なるエネルギー代謝を行っているという現象」の文中の「異なるエネルギー代謝を行っている」のは正しいのです。「異なるエネルギー代謝を行っている」のはherpesの分裂・増殖のために行われている必要な異なるエネルギー代謝なのです。
この論文の結論から先に述べると癌細胞が異なるエネルギー代謝を行なわざるを得ないのは「ヘルペスウイルスの無限大の増殖」のためにヘルぺスが、人体の細胞から奪い取る莫大なATP以外の様々な代謝性要求を満たすために間違ってワールブルグ効果といわれるあり得ない現象を高名なワーブルグが間違って見つけたのです。癌細胞が増えるために「ワーブルグ効果」が生まれたわけではないのです。
実はワールブルグが100年前に発見した「ワールブルグ効果」などは100%存在しないのです。異なるエネルギー代謝を行なわざるを得ないもうひとつ追加の説明があります。つまり正常な細胞をがん細胞に変異させるだけではなく、すでに正常な細胞を癌細胞に仕立てた後にすでに出来上がった一つの癌細胞の約800もある癌関連遺伝子もできる限り多くを癌化遺伝子に変えて癌細胞を利用できる限界まで分裂させてそのたびごとにヘルペス自身の子孫であるヘルペスビリオンもどんどんが増やすためにヘルペスは莫大な栄養素をはじめその栄養素を利用してヘルペスビリオンの完全な粒子の部品のすべてのセットをビリオンが増える度に繰り返し作るのに莫大なATPが必要となるのです。Herpesビリオンをできる限り多く生み出し、かつ癌細胞が増える方法は二つあります。一つは正常な細胞にヘルペスが感染して新たなる癌細胞にならせることです。二つ目はすでに癌細胞になった癌細胞が増殖するたびに約800もある癌関連遺伝子の残りを癌化させることです。
一個のがん細胞が最大限、増殖するには、一個の癌細胞が持っている800もある癌関連遺伝子が癌化遺伝子になればなるほど多くなればなるほど其れだけ1個の癌細胞が増殖するスピードが増えます。このようにすべての癌細胞にはATPはもちろんですが、DNA、タンパク質を構成するアミノ酸、細胞膜を構成するリン脂質、脂肪酸もたくさん必要ですこれをヘルペスウイルスが生きている限り細胞から奪い取ることをヘルペスによる代謝性要求といいます。
例えば細胞膜を構成する重要な脂肪酸であるパルミテートを合成するには、7分子のATP、16の炭素骨格、28電子(NADPH14分子)が必要です。グルコース1分子は、酸化的リン酸化により36分子のATPを生み出しますが、これではATPだけが過剰になり、NADPHや炭素骨格が足りなくなってしまいます。そこで、そこで、解糖系を回し、そこから枝分かれしたペントースリン酸経路(グルコース1分子からATP30分子とNADPH2分子を生成)も利用することで、必要なNADPH等を得るのです。
飽和脂肪酸であるPalmitate(パルミチン酸)と同様に、アミノ酸やヌクレオチドの合成においても、一見「非効率」な代謝を利用することで、がん細胞が自らの増殖/分裂に必要な炭素骨格やNADPHをバランスよく得ているのです。
興味深いことに、これらの代謝は緊縮応答ではなく、いわゆるがん遺伝子や、p53のようながん抑制遺伝子が、積極的にメタボロームを調節している結果であることが最新の研究で明らかになってきました。メタボロームとは、生体内に存在する数千種類に及ぶ代謝物質(メタボライト)の総体を指し、その種類や濃度を網羅的に分析する手法をメタボローム解析(メタボロミクス)といいます。
メタボロームとは、核酸(DNA)やタンパク質、糖、有機酸、アミノ酸などの低分子化合物が酵素などの働きによって作り出されたもので、数千種類に及びます。食事や癌などの疾病などの外部刺激によって代謝が変化すると、細胞内の代謝物質の種類や濃度にも変化が生じます。つまり癌患者の体にはヘルペスによる癌細胞、正常細胞、ヘルペスウイルス感染細胞などさまざまな細胞が入り乱れ数千種類に及ぶ低分子化合物を作るエネルギーであるATPや原料を様々な形でヘルペスビリオンを作るためにかすめ取られているのです。
例えば、癌細胞の増殖因子刺激で活性化するAkt/PI3Kシグナリングが解糖系を亢進させることや、ヘルペスのよる癌が増殖するためのチロシンキナーゼシグナルが解糖系の律速ステップを負に制御し、解糖系の中間体をペントースリン酸経路でのNADPH生成に利用させたり、反対に、がん抑制遺伝子として有名なp53がペントースリン酸経路を負に制御し、癌細胞の代謝性要求を満たす「増殖用代謝 (proliferative metabolism)」をシャットダウンしているといった体内代謝の異常を列挙するのに、枚挙に暇がないのは「癌」は特別な病気ではなく「ヘルペスビリオンが過剰に増殖する」時に生ずる「過剰すぎるエネルギー代謝」と「過剰すぎる物質代謝」が異常になりすぎた結果、最後は悪液質で死ぬのであって「癌で死ぬわけではないのです。」
Akt/PI3Kシグナル伝達経路は何でしょうか?p53などのアポトーシス作用ができないようにして癌細胞を「遺伝子による計画された細胞死」を止めてしまうのです。癌細胞や正常な細胞の増殖、生存、成長、代謝を調節する重要な細胞内シグナル伝達経路で、次のような役割を担っています。
- 正常な細胞の増殖とせいじょうな細胞死のバランスを制御することができる生存因子のシグナル伝達経路の開始点です。
- 細胞の脱分化や初期化を促進することでアポトーシスを抑制します。
アポトーシスの阻害剤として、多くの細胞生存経路において重要な調節的役割を果たします。
- 生存のために血管新生およびエネルギー代謝と物質代謝において生存のための調節的役割を果たします。
Akt/PI3Kシグナル伝達経路は、様々な成長因子やホルモンなどの細胞外からのシグナル因子からの刺激によって活性化されます。この経路の各タンパク質は、PI3K(ホスファチジルイノシトール3キナーゼ)、AKT(プロテインキナーゼB)、mTOR(mammalian target of rapamycin)で、順に活性化されます。
AKT(プロテインキナーゼB)は、細胞の増殖や代謝、生存などの基本的なプロセスを制御するセリン/スレオニンキナーゼの一種です。細胞シグナル伝達経路において重要な役割を果たし、ヘルペスが起こす糖尿病、がんや神経変性など多くの疾患に関連しています。
mTOR(mammalian target of rapamycin)は、細胞内の栄養状態やエネルギーレベル、ストレスなどの情報を感知して、細胞の増殖や代謝を調節するリン酸化酵素です。
Akt/PI3Kシグナル伝達経路の遺伝子変異や発現異常は、ヘルペスが起こすすい臓がん、白血病、前立腺がんなどの悪性腫瘍の発症に深く関与しています。
ワールブルグ効果とは何であり本当に存在するのでしょうか?実はないのです。
ワールブルグ効果とは、高名なドイツの生理学者であるオットー・ワールブルグ博士が発見した、「がん細胞が正常細胞とは異なるエネルギー代謝を行っているという現象」です。もっと具体的に言えばワーブルグ効果では、酸素が十分に存在している条件下でもミトコンドリアの活性を抑え、グルコース代謝を主に解糖系にシフトさせます。ドイツの生理学者であるオットー・ワールブルグ博士は,がん細胞が正常細胞とは異なるエネルギー代謝を行っていることを発見したことになっています。何故ワールブルグ博士は,がん細胞が正常細胞とは異なるエネルギー代謝を行っているという間違いを犯したのでしょうか?がん細胞はヘルペスウイルスが細胞に感染してその細胞の代謝を変えてしまったためであることに気が付いていなかったからです。勿論現代の癌学者さえ気が付いていないことですから100年前にヘルペスウイルスが遺伝子を癌化させていることをワールブルグ博士の様な偉い医学者も知る由もないからです。
酸素が十分に存在している条件下でも,ミトコンドリア活性を抑え,主に解糖系にグルコース代謝をシフトさせるという特性は「ワールブルグ効果」として認知され,ワールブルグ効果によるがん進展説は今もなお有力な作業仮説であるのですがこの作業仮説100%間違いであることを誰も証明できないのです。作業仮説とは、研究や実験の過程で、十分に実証されたものではないものの暫定的に有効とみなされる仮説です。
しかしながら,ワールブルグ効果による癌進展説はいまだにいくつかの点が解決されていないのです。たとえば,ワールブルグ効果は①発がんの直接的なドライバー因子(発がんに直接関係する遺伝子)になりうるのか,②もしくは悪性化した進行したがん細胞の増殖や浸潤に必要な二次的な性状変化なのかについては,いまだ明確な答えが得られていない.勿論一般的にワールブルグ効果はがん細胞の生存・増殖を助長すると考えられているが,これも誤りなのです。がんの各ステージにおける腫瘍促進作用は正確にはわかっていないのも誤りなのです。世界中でヘルペスが癌のしっているのは「松本漢方クリニック」の15歳からあらゆるヘルペスによる難病で苦しみ続けて最後は医者になってすべての病気の原因はヘルペスであることに気づいた松本院長だけなのです。「癌とは殺し切ることができない無限に増殖し続けるヘルペス感染細胞のヘルペスによる異常なエネルギーかつ物質代謝に起こされた病気に過ぎないのです。更に別の言い方をすれば「ゲノム内感染ヘルペスによる増殖遺伝子の変異」に過ぎないのです。にもかかわらず、この「ワーブルグ効果」を基にして癌はミトコンドリアの機能障害が原因であるとか活性酸素、放射能、紫外線などによって傷ついたDNAの蓄積によって癌増殖関連遺伝子が癌化したためだと言われたりします。このふたつの理論は完全に間違っていることを証明していきます。さらに本当の癌の原因は「ゲノム内に感染したヘルペスによる増殖遺伝子の変異した細胞の過剰増殖である」ことを証明します。
ワールブルグ効果とミトコンドリアと癌の関係は、一般的には①酸素が十分に存在している条件下でも、ワールブルグ効果によりミトコンドリアの活性を抑える。②その結果グルコース代謝を主に解糖系にシフトさせる。③ワールブルグ効果は、がん代謝の分野で注目を集めてきた腫瘍形質(腫瘍遺伝子)のひとつとされています。④腫瘍形質(腫瘍遺伝子)を発現したがん細胞は、酸化的リン酸化と比較してグルコースからATPを産出する能力が著しく低下しているにもかかわらず、好気的解糖を続けているとされています。⑤ワールブルグ効果の意義はじつはよくわかっていないという問題を秘めています。⑥人によってワールブルグ効果の定義がまちまちだったり誤解も多く見られます。
ここでワールブルグ効果について,PKM(Pyruvate kinase M)ノックインモデルを通してもう一度見つめなおします。解糖系酵素M(Pyruvate kinase M; PKM)とは解糖系酵素であるピルビン酸キナーゼM(Pyruvate kinase M;PKM)です。解糖系酵素であるピルビン酸キナーゼM( PKM)には高活性型のPKM1と低活性型のPKM2という2つのスプライシングisoformが存在し,大半のがんがPKM2を選択的に発現する。かつては,PKM2はワールブルグ効果との関連が深く腫瘍に代謝上の有利をもたらすとされていたのですが,PKM2欠損マウスの表現型は“がん促進”だったのです。PKM2が働かない時の方が癌が進行するのです。
「ワールブルグ効果」は,がん代謝の象徴ともいえる現象であるが,その意義はよくわかっていない。人によって「ワールブルグ効果」の定義がまちまちだったり誤解が多いという問題もあります。さらにワールブルグ効果とは、高名すぎるノーベル賞受賞者であるドイツの生理学者のオットー・ワールブルグ博士が100年前に発見した長い歴史のゆえかもしれないのでいまもなお正しいと信ずる研究者も数多くいます。
今回,この癌の代謝形質と密接に関連する解糖系酵素PKMの新たな遺伝子改変マウスモデルを用いた解析から,ワールブルグ効果が,腫瘍細胞にとって,むしろ代謝上のハンデ(不利)となっている可能性が浮上してきたのです。代謝形質とは、代謝に関する表現形質を指します。まず代謝とは、生物の体内や細胞内で起こる化学反応のことで、生命維持のために生体内で化合物を合成する一連の反応です。食事や呼吸によって取り入れた栄養や酸素を基に、生体物質やエネルギーを生み出します。一方、表現形質とは、生物個体の形や性質を指し、遺伝子によって決まる特徴です。たとえば、人間の身長は、成長や食欲、筋肉量、活動レベルに影響を与える遺伝子など、多くの遺伝子によって決定されます。実際,一部の高悪性肺がんでは,PKMの発現アイソフォームがアンチワールブルグ効果型になっています。最近,新たなPkm遺伝子改変マウス群を作製し,がんや代謝制御におけるPKM1・PKM2の役割を再検証されました。マウス発がん試験や細胞移植実験などを通じ,真に腫瘍促進的なのは,PKM2ではなく,驚いたことにPKM1の方であることがわかったのです。実際にPKM1を高発現し,その生存・増殖を腫瘍に代謝上の不利をもたらすとされていたPKM1に依存する高悪性がんの存在も明らかになったのです。
1. ワールブルグ効果とは何か?
ワールブルグ効果(Warburg effect)は,がん代謝の分野において,これまで最も注目を集めてきた腫瘍形質(癌の遺伝的形質)です。言い換えると腫瘍形質とは癌細胞の核の中に存在する遺伝子または遺伝子群によって表現形質である癌になります。遺伝子はDNA(デオキシリボ核酸)を構成するアデニン(A)、グアニン(G)、シトシン(C)、チミン(T)の4種の塩基の配列順序によって決定されています。
ほぼ1世紀前,O. Warburgらは,当時提唱されていたパスツール効果が腫瘍組織においてはみられないことを観察しました(図1)。すなわち,正常組織とは異なり,腫瘍組織においては,“酸素分圧を上げてもグルコース依存性の乳酸産生が下がらない”ことを発見した。パスツール効果とは細胞や微生物やウイルスよる解糖や発酵が酸素によって抑制される現象です。フランスの細菌学者であるパスツールが発見したことに由来しています。パスツール効果の仕組みとは①酸素濃度が十分な場合、細胞は好気的な条件でATPを消費することで大きなエネルギーを得ることができます。②細胞が必要とするエネルギー量はほぼ一定なので、ATP生成量が多い呼吸が行われている場合、解糖は抑制されても差し支えないと考えられます。③この作用は、グルコースやグリコーゲンなどのエネルギー源の無駄な消費を防ぐ働きであると考えられています。④パスツール効果は、酵母の発酵についていわれていましたが、現在は動物についても使われています。ところが腫瘍組織では,酸素濃度が十分に高い条件でも,TCA回路の回転(CO2の生成)と比較した際,乳酸産生量が高い傾向がある,ということなのです。ワールブルグ効果とはそのような内容と定義して話として進められるのです。

(ア)検圧計の模式図.Warburgが用いたワールブルクの検圧計(Warburg’s manometer)は生物材料のガス代謝を測定するのに用いられる一種の圧力計。U字管の先端に反応用容器を取付け,容器内のガスの出入りを,U字管両脚内の液面の差として読取る。検圧計の模式図.S:組織スライス,R:リンゲル液,C:吸収剤(KOH等)チャンバー,P:基質ポート(出入口).(イ)グルコース代謝模式図.
Warburgの観察とそれにまつわる考察が,その後のがん代謝研究の進展に与えた影響はきわめて大きい.しかし,同時に,主に技術的な制約から,実験の条件設定や解釈において問題がなかったわけではない.そのため,少し考えてみると,現代のがん代謝モデルにそのまま当てはめるのが容易ではないことに気づく.たとえば,Warburgらは組織スライスの代謝活性をモニターしたわけだが,その際,組織が浸されたリンゲル液には,グルコース以外の炭素源が含まれていない.腫瘍はグルコース以外にも多くの栄養素を炭素源として利用していることを,現代の我々は知っている.グルタミンやアミノ酸,より最近では,これまで主に単なる排泄物と考えられていた乳酸までもが,がんにとって重要な炭素源となっていることが明らかになっている。当然のことながら,Warburgの計算において,一度排出された乳酸が再利用されるなどという可能性は考慮されていない.また,冒頭において,グルコース“依存性”の乳酸産生とわざわざ記したのは,その乳酸が本当にグルコースに由来しているという確証がないからだ。培地中にグルコースを添加すると,乳酸産生が,一気に,そして劇的に高まるのは確かである.しかし,その際,新たな乳酸生成にグルコース由来の炭素ばかりが使われているかというと,答えは否だ.
このように,彼らの結果・解釈にはさまざまな問題が内包されているが,中でも,好気呼吸について,見過ごせない大きな誤解・齟齬がある.Warburgらは,冒頭に述べたような観察結果を,“腫瘍細胞ではグルコースの酸化(TCA回路)が、正常組織と比べて抑制されているため”と解釈した.そしてその原因を,好気呼吸の場であるミトコンドリアの異常に求めた。ほとんどの癌で腫瘍細胞の好気呼吸システム自体に異常がないことは広く知れわたってきている。しかし,問題はそこではない.
新たな技術(安定同位体トレーサー解析や細胞外フラックス解析装置など)の普及に伴い,“腫瘍の好気呼吸レベルは、対応する正常組織と比べて低い”という推論がそもそも正確ではないことが近年明確になってきている。少なくとも米国での動向をみると,その部分の誤解はずいぶん解消されたと感じる.以前よく見かけた“がん代謝”のモデルでは,エネルギー代謝が,正常組織での好気呼吸から,がんでの解糖系へと「スイッチ」する(旧モデル,図2).これに対し,最近の理解は,「がんでは解糖系・好気呼吸の両方の活性が,正常組織・細胞と比べて上昇している」というモデルへと改められた(新モデル,図2).この意味は「癌」はヘルペスビリオンが大量に増殖することですからこのヘルペス増殖のときに代謝エネルギーがグーンと増える必要があるので、癌では解糖系・好気呼吸の両方の活性が,正常組織・細胞と比べて上昇するのは当然の「理」なのです。そうでなければ正常細胞が癌になることはあり得ないのです。
がんと正常組織を比べた場合,好気呼吸の活性化度合いは解糖系のそれより低いかもしれないが、しかし,がんの好気呼吸が正常組織・細胞よりも低いというのは明らかに誤り,というのが現代の世界の趨勢なのに、日本では多くの研究者たちが「がんではエネルギー代謝が好気呼吸から解糖系へと切り替わる」とか「正常と比べ,がんではミトコンドリア好気呼吸が抑制されている」と断言しているのは二つの間違いをおかしているのです。一つ目はワールブルグ効果が今なお正しい。という間違いと、二つ目はいまだに世界や日本の癌学会においてはすべての癌はヘルペスウイルスによるヘルペスが増え続けるために癌関連遺伝子(正しくは増殖遺伝子)がherpes自身の遺伝子によって変異してしまった「ヘルペス性の細胞異常増殖症である」という真実に気が付いていないのです。

図2 がんにおける解糖系と酸化的リン酸化の関係
正常組織との比較において,がんでは,酸化的リン酸化(OxPhos)から解糖系へとスイッチするモデル(旧モデル)から,解糖系・酸化的リン酸化の両方が亢進しているというモデル(新モデル)へと,認識が改められた。OXPHOS(酸化的リン酸化)とは、細胞呼吸(糖代謝)の最後の段階でミトコンドリアの内膜で行われる一連の酸化還元反応です。この反応において、ミトコンドリア内膜の5つの膜貫通型酵素複合体が電子を1つの分子から別の分子に移動させ、ADPと無機リン酸からATPを合成して細胞のエネルギーの大部分を生み出します。
ミトコンドリアは「細胞のエネルギー発電所(powerhouse)」とも呼ばれ、エネルギー産生以外にも数多くの代謝反応の場として細胞機能を維持する上できわめて重要な役割を果たしています。.
2. ピルビン酸キナーゼM(Pyruvate kinase M;PKM)とワールブルグ効果について。ワールブルグ効果のメカニズムと問題点は何か?
さて,あれこれ問題があるワールブルグ効果であるが,多くのがんが,そのような代謝形質(高い乳酸産生/好気呼吸比)を示すのは事実である.しかし,この乳酸産生の方が好気呼吸よりも多いという代謝形質のがんにおける役割はよくわかっていない.この点において,この10年ほど,強い関心を集めてきたのが解糖系酵素であるピルビン酸キナーゼM(PKM)である.PKMは,解糖系における三つの不可逆反応のうちの一つ,ホスホエノールピルビン酸からピルビン酸への反応を触媒する.ピルビン酸は,TCA回路に動員される可能性と乳酸へと代謝される可能性の両方がある,代謝ネットワーク上でハブ(中心)となっている代謝物である(図3).

図3 グルコース代謝とPKM
さて,あれこれ問題があるワールブルグ効果であるが,多くのがんが,そのような代謝形質(高い乳酸産生/好気呼吸比)を示すのは事実である.しかし,この乳酸産生の方が好気呼吸よりも多いという代謝形質のがんにおける役割はよくわかっていない.この疑問に対して答えを出しましょう。癌になってしまった時ほど人生の中で最も強いストレスをかんじることはありません。死ぬか生きるかのストレスほど恐ろしいストレスは他にありません。
しかも免疫が最高に落下する時でもあります。癌の原因はヘルペスウイルスですから癌になる前にストレスによって8種類のherpesの感染が多かれ少なかれ生じています。癌と宣告されれば最も免疫がさらに低下してしまいます。この時に感染細胞のゲノムに潜伏感染していたよるヘルペスウイルスの仲間たちが癌細胞や正常細胞が分裂増殖するときにつまり一般的には再活性化する時に、同時にヘルペスも活性化して分裂して増え出しヘルペスビリオンを莫大な数と言えるくらいに大量に作り出します。
この増殖のときに作り出すすべての完全なherpesウイルスビリオンの構造蛋白であるスパイク、エンベロープ、テグメント、カプシドなどに加えて、ヘルペスウイルスのDNAやDNAの材料になる塩基や多種類のグリコプロテインなどの部品を作り続けるのにどれだけの原料とエネルギーを細胞がヘルペスに提供せざるを得ないのかを想像できますか?
細胞が分裂する時にのみヘルペスも分裂できるのでherpesを再活性化(分裂)させる誘導の機序に関しては,ストレスによって免疫力が低下すること以外に,細胞を分裂させる活性化因子も必要でインターロイキン 6(IL-6),インターフェロ ンと,ケモカイン類などの免疫のサイトカインも働いているのです。
乳酸は、筋肉を動かすためにエネルギーを得る際に生成される物質で、筋肉に蓄えられたグリコーゲンを分解する必要があります。このグリコーゲンの分解の過程で、ピルビン酸が乳酸に分解されます。このエネルギー発生の仕組みは乳酸性機構と呼ばれます。このグリコーゲンの分解の過程で、ピルビン酸が乳酸に分解することでエネルギーが発生します。このエネルギー発生の仕組みは乳酸性機構と呼ばれます。
糖質が解糖系(嫌気的代謝)で分解されることで作られます。乳酸が生成される仕組みは次のとおりです。
癌患者も生きて筋肉を動かすために、筋肉に蓄えられたグリコーゲンを分解する必要があります。このグリコーゲンの分解の過程で、ピルビン酸が乳酸に分解されます。このエネルギー発生の仕組みは乳酸性機構と呼ばれます。乳酸性機構とは、筋肉でエネルギーを作るときに糖質が解糖系(嫌気的代謝)で代謝・分解されて乳酸が生成される過程を指します。
癌患者の筋肉を収縮させるエネルギーを得るために、筋肉に蓄えられたグリコーゲンをピルビン酸から乳酸に分解することでエネルギーが発生します。
乳酸は運動をした後に蓄積することから疲労物質とも呼ばれてきましたが、乳酸が作られる過程で発生する水素イオンなどの作用で、筋肉のpHバランスが酸性に傾くことが疲労の一因と考えられています。癌患者は常に活動するとすぐに疲労困憊する原因は筋肉のpHバランスが酸性に傾くからです。
しかし、乳酸は疲労物質どころか、癌患者の筋肉、脳、心臓のエネルギー源となる大切な物質であるのです。また、乳酸には癌患者の血管新生や傷の修復促進、ミトコンドリア新生、遺伝子発現調節などの働きもあります。しかも乳酸は無酸素状態になると作られるエネルギー源になる物質です。無酸素状態になると作られた乳酸をエネルギーに変えて利用するために、軽い運動で血流を良くして乳酸を血液で肝臓まで運ばせ、肝臓でグリコーゲンに再合成され、再びエネルギー源であるグルコースとして再利用させることができるのです。
グリコーゲンの代謝はどうなっているのでしょうか?グリコーゲンは,グルコースがα-1,4グリコシド結合で重合した直鎖構造と,α-1,6グリコシド結合によって枝分かれした構造が組み合わさったものであり,グルコースの貯蔵体である。グリコーゲンは,肝臓には肝臓の重量の約5%(約100 g),筋肉には筋肉の重量の1%(約250 g)が含まれている。肝臓内のグリコーゲンは分解されてグルコースとなり,主として空腹時の血糖値を維持するための原料である.筋肉内のグリコーゲンは,運動をする際のエネルギー源であり,癌患者の解糖系で作ったグリコーゲンを癌患者も生きている間は筋肉を動かすために、筋肉に蓄えられたグリコーゲンを筋肉内で分解されて解糖系を経て筋収縮に必要なATPを産生します。この過程は「解糖」と呼ばれ、細胞質で行われる嫌気的な反応です。解糖の過程では、グルコースがピルビン酸に分解される。ピルビン酸と水素が結合して乳酸が生成される
このグリコーゲンの分解の過程で、ピルビン酸が乳酸に分解することでエネルギーが発生します。このエネルギー発生の仕組みが乳酸性機構なのです。筋肉でエネルギーを作るときに、グリコーゲンが分解されてピルビン酸が乳酸に分解されます。
乳酸性機構とは、筋肉でエネルギーを作るときに糖質が解糖系(嫌気的代謝)で代謝・分解されて乳酸が生成される過程を指します。
乳酸は、筋肉を収縮させるエネルギーを得るために、筋肉に蓄えられたグリコーゲンをピルビン酸から乳酸に分解することで生成されます。
ピルビン酸がアセチルCoAとなり有酸素運動で消費されない場合、ピルビン酸は乳酸に分解されます。 このエネルギー発生のしくみを乳酸性機構といいます。乳酸は、筋肉でエネルギーを作るときに糖(グリコーゲン)が分解されて生成されます。筋肉中に蓄積された乳酸は血液によって肝臓に運ばれ、休息などによって酸素が補給されると、乳酸の大部分はグリコーゲンに再合成され、ATPが合成されます。グリコーゲンから肝臓でATPが作られる経路は、無酸素性エネルギー代謝の解糖系です。
解糖系では、グリコーゲンやグルコースを分解することでATPが産生されます。グリコーゲンは、肝臓グリコーゲンとして貯蔵されており、血糖を一定に保ち、空腹時の糖質のグルコースを供給源として機能しています。
グリコーゲンの分解は、グリコーゲンホスホリラーゼの作用によってグルコース 1-リン酸が切り出され、一分子短いグリコーゲンとなります。グルコース 1-リン酸は、グルコース 6-リン酸へと転換され、肝臓ではグルコース-6-ホスファターゼによってグルコースに変えられ、血中に放出されます。
グリコーゲンが蓄えられるのは、主に筋肉と肝臓であるが両者のスタンスは全く異なる。筋肉は蓄えたグリコーゲンをすべて自分で消費して他に分け与えることはしない。これに対して、肝臓はもっぱら他の組織や器官に配給するためにグリコーゲンを蓄え、必要に応じて血液中に放出する。食事と食事の間でも血液中のグルコース濃度が一定レベルに維持できるのはこのためです。
乳酸性機構とは、ピルビン酸がアセチルCoAとなり有酸素運動で消費されるのですが、消費されない場合、ピルビン酸は乳酸に分解されます。筋肉でエネルギーを作るときに糖質が解糖系(嫌気的代謝)で代謝・分解されて乳酸が生成される過程を乳酸性機構といいます。
筋肉を収縮させるエネルギーを得るために、筋肉に蓄えられたグリコーゲンをピルビン酸から乳酸に分解することで乳酸は生成されます。
また、乳酸は筋肉からカリウムが漏れ出して筋収縮を阻害することを防ぐ働きがあります。
この乳酸性機構が多くの癌で,癌に特有なエネルギー代謝であるそのような代謝形質である「高い乳酸産生/好気呼吸比」を示し高い乳酸産生を行っているのです。癌細胞が分裂するたびごとにherpesビリオンを大量に増殖させるために余計な厖大なエネルギーを使うので乳酸性機構が活発に働きだした結果、この乳酸産生の方が好気呼吸よりも多いという代謝形質の癌における役割がエネルギー産生のために活発に働きだしたのです。
2008年,この酵素の性質とワールブルグ効果との密接な関連を指摘する,非常に重要な報告がなされた。具体的には,PKMには選択的スプライシングで生じる二つのアイソフォーム(PKM1, PKM2)が存在するが,これらのうち,がんにおけるPKM2の選択的発現が,腫瘍細胞のワールブルグ効果に非常に大きく寄与していることが明らかにされた(図4).これまで知られるほぼすべてのがんは,PKM1ではなく,PKM2を主に発現しているのだが,それをPKM1で置き換えると,ワールブルグ効果がキャンセルされることが示された.

図4 PKMのスプライシングアイソフォームとワールブルグ効果
(ア)PKM1とPKM2のバランスは,ワールブルグ効果の成立・解消を制御する.(イ)PKM1は構成的な高活性型,PKM2は条件的な活性化型である.
それでは,なぜ,ワールブルグ効果形成のためにはPKM1ではダメなのか? その原因は,両アイソフォームの活性制御の違いが重要とされている(図4).PKMは,四量体を形成することで初めてその酵素活性が現出する.PKM1は,特に他因子に依存することなく,発現すると自動的に四量体を形成する.つまり,PKM1は,構成的な高活性型である.一方,PKM2は,通常,単量体あるいは二量体として細胞内に存在する.PKM2が四量体を形成するには,アロステリック活性化因子(F1, 6BPやセリンなど)の存在が必須であり,そのため,それら因子の細胞内レベルが上昇したときのみ活性化する.アロステリックとは、酵素や受容体などのタンパク質の活性部位やリガンド結合部位以外の部位(アロステリックサイト)にエフェクターと呼ばれる化合物が結合することで、タンパク質の活性や機能が調整される現象を指します。アロステリックという言葉は、ギリシャ語で「別の(allo-)」と「形(stereo-)」を意味する合成語です。アロステリックサイトに結合するエフェクターは、アロステリックモジュレーターと呼ばれ、シグナルの強度を高めるポジティブアロステリックモジュレーターと、シグナルの強度を弱めるネガティブアロステリックモジュレーターに大別されます。モジュレーター(modulator)とは英英語の元の意味は「調節器」ですが
代謝活動に関する酵素については、その酵素の本来の基質と全く異なった基質化合物が酵素の活性部位以外に結合することにより、酵素活性が変化するものがある。このような酵素をアロステリック酵素といい、これに結合してその酵素活性を調節する化合物をモジュレーターという。つまり,PKM2は,条件的な活性化型である.そのようなアロステリック制御は,結果として,PKM1と比べた場合,細胞におけるピルビン酸の生成レベルを低く保つことになる.そのような作用を通じてPKM2がもたらす低い“グルコースからピルビン酸へのフラックス(流れ)”が,そのまま下流であるTCA回路への炭素源流入を制限するモデルが提唱された。この際,PKM2によってピルビン酸生成が低下する一方,なぜ同時に乳酸産生の亢進が起きるのか?という点は謎とされ,そのまま現在に至っている.
もう一点,以前,“正常細胞はPKM1型、がんはPKM2発現型”というモデルがあったが,少なくない研究から,これは誤りであることが明確になっている.大半の上皮細胞を含め,がんの起源となるような細胞のほとんどは,いかなる発がんイベントもなしに,元来PKM2を発現している.正常細胞におけるPKMのアイソフォーム選択は,主に,細胞の分化系列によって規定されることが示唆されている.一方で,ほぼすべての腫瘍細胞がPKM1よりもPKM2を圧倒的に発現していることは確かであり,この事実は,多くの腫瘍がワールブルグ効果を示すことと一致している.それでは,PKM2(および,それがもたらすワールブルグ効果)は,がんを促進する作用があるのか?
当初,いくつかの細胞株を用いた実験成績は,上記アイディアを支持するものだった.ところが,PKM2特異的なノックアウト(KO)マウスが作製され,その表現型が明らかになるにつれ,事態は一気に混沌化することになる. PKM2-KOマウスは,PKM遺伝子にて,PKM2特異的なエキソンのみを欠失させた遺伝子改変マウスである.驚いたことに,そのようなPKM2欠損マウスにおいて,がんが促進されることがわかった.PKM2は,がん促進的どころか,がん抑制的である可能性すら出てきたのである.しかしながら,PKM2-KOマウスは,その問題を扱うには不向きな,あるシステム上の問題を抱えていた.具体的には,PKM2欠損に伴って,代償的に,PKM1が中途半端な程度に発現するという問題である(図5).そのため,PKM2-KOは細胞のPKM活性を低下させているのか,それとも上昇させているのか判然としないことになってしまった.また,Cre-loxPシステムと併用した場合には,その組換え効率が100%ではないことも,よりいっそう,結果の解釈を困難にした。
Cre-loxPシステムとは何でしょうか?Cre/loxPシステムは、DNA組換え酵素CreとloxP配列と呼ばれるDNA配列を利用した遺伝子組換え実験系です。DNA組換え酵素CreとloxP配列とはDNA組換え酵素CreとloxP配列は、遺伝子組換え実験系であるCre/loxPシステムにおいて相互作用することで、部位特異的な組換え反応を起こします。loxP(ロックスピー)配列とは、特定のDNA配列で、DNA組換え酵素Creが働くことで部位特異的組換え反応を起こす配列です。loxPは元来バクテリオファージP1のゲノム中にある34塩基からなる配列であり、 中央8 bpは非対称な配列でその両側13 bpが対称になっている。 loxはlocus of crossing overに、PはファージP1に由来している。loxP配列は、8塩基対のコア配列スペーサー領域で区切られた2つの13塩基対のインバーテッドかつパリンドローム・リピート(逆位反復配列)から構成されています。インバーテッドとは 「逆転の」「逆位の」という意味でパリンドロームとは「回分の」の意味で、「回分」とは「前後どちらから読んでも同じ語句」という意味です。バクテリオファージP1とは、大腸菌に感染する溶原性ファージで、ホストDNAの一部を取り込み、形質転換を起こす。レプリコンを取り出してプラスミド化できる。レプリコンとは「レプリカ(複製)」という言葉から造られた分子生物学の用語で「コピーされたDNAもしくはRNA」を意味します。レプリコンワクチンとはこれまで新型コロナウイルスに対して使用されてきたmRNAワクチンは、体内にウイルスのタンパク質をつくることで抗体や免疫を獲得しますが、そのタンパク質はそのまま消滅します。一方でレプリコンワクチンはmRNA自体を複製する酵素も組み込まれており、ウイルスのタンパク質を体内で作り続けます。この「自己増幅」によって従来のmRNAワクチンよりも強い免疫反応を長期間獲得できるとされている自己増殖型のmRNAワクチンです。 体内で自己増殖することで、少量のワクチンでも長期間にわたって免疫効果を持続させることが期待されていますが問題もあります。ファージP1とはファージP1によってコードされるCREタンパク質は、大腸菌とin vitro(試験官内で)の両方で分子内および分子間シナプスとDNAの再結合を促進する。ファージの基本構造は、タンパク質の外殻と遺伝情報を担う環状の核酸(主に二本鎖DNA)で構成されています。細菌にとりつき、細胞膜に穴を開けて内部で増殖し、細菌を死滅させるものが多いです。herpesウイルスと非常によく似ているのでだれも明らかにしていないヘルペスのゲノムをヘルペスがヒトのゲノムのDNAに組み込んでヒトのゲノムのDNAを組み替えているかの答えを出そうと努めているのです。もちろん異なる点もあることは言うまでもないことです。
ファージ(バクテリオファージ)は、宿主細菌のいるところどこにでも存在しており、地球には1031個以上存在するのでファージ(バクテリオファージ)は地球全生物の総数より多いのです。
バクテリオファージP1のゲノムには、loxPと呼ばれる34塩基からなる配列があります。loxPは、locus of crossing over(交差部位)とファージP1に由来しています。loxP配列は、DNA組換え酵素Creが働くことにより部位特異的組換え反応を起こします。これは、バクテリオファージP1が宿主である大腸菌内で複製される際に自身のゲノムを環状化するための組換えシステムです。
下図のPA-CREはPA-Cre(光活性化型Cre)とは、光照射によってDNA組換え反応を制御できるシステムです。遺伝子の働きを時間や空間特異的にコントロールすることで、遺伝子の生体機能を解明するモデルリソースとして期待されています。
l
Creリコンビナーゼは、細胞内の遺伝子やLoxPの DNAのシーケンスを識別して組換え反応を起こす酵素で、コンディショナル遺伝子ノックアウトマウスの作製に必要となります。DNAシーケンスとはDNAシーケンスとは、DNA(デオキシリボ核酸)を構成するヌクレオチドの配列(塩基配列)を決定するプロセスです。DNAは生物の遺伝情報を担う分子で、その塩基配列を調べることで遺伝情報を解析することができます。コンディショナル遺伝子ノックアウトマウスとは、特定の遺伝子の働きを任意の時期や場所(臓器など)で機能しなくするように工夫したマウスで条件特異的遺伝子破壊とも呼ばれ、遺伝子機能を解析するのに有用な方法です。遺伝子機能を解析する実験動物として広く利用されており、遺伝子機能を解析するのに非常に有用な方法となっています。コンディショナル遺伝子ノックアウトマウスの作製には、Cre/loxPシステムが使用されることが多く、その原理は次のとおりです。①標的遺伝子にloxP配列を挿入する②特定の時期や場所でCre組換え酵素を発現させる③Cre組換え酵素がloxP配列の両端の逆位反復配列と結合して二量体を形成する④二量体は他のloxP部位の二量体と結合して四量体を形成する⑤loxP部位の間のDNA配列はCre組換え酵素によって切り落とされ、切り口はDNA連結酵素によって再接続されるのです。
Creリコンビナーゼとは、バクテリオファージP1に存在するリコンビナーゼです。Creリコンビナーゼは、34塩基からなるDNA配列(loxP)に結合して部位特異的に組換えを起こします。
コンディショナルノックアウトは、条件特異的遺伝子破壊とも呼ばれ、遺伝子機能を解析するのに有用な方法です。Floxマウスとは、 欠損させたい遺伝子の両端にそれぞれ34塩基からなるlox配列が組み込まれたマウス。 loxに挟まれた領域は、DNA組換え酵素(Cre)により組換えが起こり除去され、遺伝子が破壊されます。Creマウスとは、即ちCreリコンビナーゼがマウスの特定組織または細胞で発現するマウスである。 そのCre遺伝子の発現は一つの特異性プロモーターに制御され、このプロモーターがCre発現の組織や細胞のタイプを決定するのです。
Cre/loxPシステムでは、細胞が酵素のCreを生産すると、その細胞のゲノム中にあるloxP部位同士の間で組換えが起きます。loxP同士の向きによって組換えの結果が異なり、次のようなことが起こります。向きが逆であれば挟まれた配列は反転する。同じ向きだと挟まれた配列が切り出される。この結果、染色体の一部を転置・欠失させたり、染色体同士を転位させたりすることが可能になります。遺伝子工学や発生工学の分野で、loxP配列間に存在する遺伝子を欠失させるために使用されています。遺伝子工学や発生工学の分野で、特定の遺伝子を欠失させる、またはノックイン(挿入)するために広く利用されています。
Cre/loxPシステムの主な特徴は①効率が高い Cre組換え酵素とloxP部位を持つDNAフラグメントが二量体を形成することで、DNA組換えが簡単で効率的に行われます。②特異性が強い loxP配列の唯一性により、遺伝子組換えの特異性を保証します。③応用範囲が広い Cre組換え酵素は、生物の異なる組織や生理条件で作用することができます。Cre/loxPシステムの利用例としては、①条件付きノックアウトマウスの作製②生体での遺伝子のはたらきを解明するための技術として用いられます。Cre/loxPシステムの利用には、次のような手順があります。一つ目は欠損させたい遺伝子の両端にlox配列を組み込んだfloxマウスを作製します。二つ目は組織特異的にCreリコンビナーゼを発現するCreマウスとfloxマウスを交配させます。三つ目はCreリコンビナーゼが発現する特定の組織のみで特定の遺伝子が欠失します。
図5 Pkm2-KO(ノックアウト)モデルとその問題点
PKM2特異的エキソンの欠損により,Pkm pre-mRNAは,その約6割が異常なスプライシングを受け,残り約4割がPkm1型となる.
Cre-loxPシステムとは何か?Cre-loxP部位特異的組換えのことです。DNAの組換えのシステムには二種類があります。DNA相同組み換えとDNA部位特異的組換えの二つです。
遺伝物質DNAの主な活動には①複製②修復③組み換えの三つがあります。この三つは密接に関係しており「修復」「組み換えは」どこかのステップで必ず「複製」をともないます。さらに「組み換え」と「修復」には共通部分がおおいのです。しかも「修復」や「組み換え」は即遺伝子に関わりがあるので「遺伝子の複製」に劣らぬくらいに生命の根源に関わっているのです。つまり修復と組換えは遺伝学の近質的な部分と関わっており「修復」は変異(突然変異)との関連が深くしかも変異(突然変異)はすべての「癌」やすべての「遺伝子病」はまさに難病である遺伝子疾患の中心を占めているのです。勿論言うまでもなく変異(突然変異)を起こすのは原核生物である単細胞から成り立っている細菌の遺伝子を簡単に変えるように多細胞で成り立っている真核生物の1個の細胞にヘルペスウイルスまず感染してその細胞のゲノムに自分のゲノムを遺伝子に組み込みをその遺伝子を簡単に組み替えてしまい「癌」や「遺伝子病」を生みだすのは正にファージ同じ「組み換え」のシステムをヘルペスウイルスが利用しているシステムそのものです。
何故遺伝子のDNA分子は修復と組換えが必要なのでしょうか?生体分子の多くは「使い捨て」です。タンパク質やRNAは遺伝情報があれば合成可能ですしそれ以外の生体分子も「生合成」で作れます。ところがDNAが損傷してしまうとスペアがないので傷がついても何とか復元する必要があるのでDNA修復はこのために生まれたのです。しかも生命は遺伝子の持つ遺伝情報で生命が維持されているのでしかも正常な遺伝子は正常なDNAによって維持されていますから長い長い進化の歴史の中であらゆる種類のDNA損傷をDNA修復酵素で事実上100%治癒できる様に進化したのです。1日1細胞当たり、1万から100万箇所の頻度でDNAが損傷を受けているといわれているのにこれが修復されずに蓄積され続ければ人類はとっくの昔に滅びてしまっているでしょう。
DNAの損傷には内因性と外因性とがあり内因性の損傷は細胞の中で日常的にできるもので生まれる損傷の数も極めて多いのです。外因性の損傷は外からの有害物質によるものですが通常の生活の中では規制が厳しいのでDNA損傷をおこす物質は過剰に摂取出来ないようになっているのですがherpesウイルスは規制しようがないのでしかも医療で用いられる薬は本当のワクチン以外は病気の原因療法のための薬剤は何一つなく免疫を抑制することのよってか症状を軽減する対症療法だけですからヘルペス天国になってしまっているので癌も増えるばかりとなってしまっているのです。
内因性であろうが外因性であろうがいかなるDNA損傷は「塩基損傷」と「DNA切断」の二つに分けられます。すでに述べたように細胞に備わったDNA修復酵素をはじめとする
DNA修復系は長い進化の中であらゆる種類のDNA損傷を事実上100%治癒できる修復システムを持っているのです。直接的なDNA損傷は「塩基損傷」と「DNA切断」の二つしかありません。DNA修復酵素とは何でしょうか?DNAを修復する酵素には、次のようなものがあります。
- 光回復酵素
紫外線によって損傷したDNAを、太陽光中の青色光成分で修復する酵素です。フラビンアデニンジヌクレオチド(FAD)と呼ばれる補酵素を持ち、光を吸収して電子を損傷DNAに移動させることでDNAを修復します。
DNAグリコシラーゼ、APエンドヌクレアーゼ、グリコシラーゼ関連APリアーゼ
塩基除去修復(BER)を行う酵素です。
- DNA Polymerase Ⅳ(Dpo 4)
損傷乗り越え複製(TLS)を行う酵素で、PCRに用いることで損傷したDNAを増幅することもできます。
- Form-amido-pyrimidine-DNA Glycosylase(Fpg)
DNAの変異部位のホスホジエステル結合を切断する酵素です。
- 8-oxoGuanine DNA Glycosylase(hOGG1)
二本鎖DNA中の8-oxo G/C、FAPY/Cの塩基対を認識し、8-oxo Gを除去する酵素です。
⑤MutY DNA Glycosylase
アデニン/グアニンまたはアデニン/8-oxo-デオキシグアニンのミスマッチ部位からアデニンを切断する酵素です。
⑤Thermostable TDG Protein
チミジン/グアニンのミスマッチからチミジン塩基を除く酵素です。
DNAは、放射線や食物の発がん物質、たばこ、環境中の化学物質、活性酸素などによって損傷を受けます。1日1細胞当たり、1万から100万箇所の頻度でDNAが損傷を受けているといわれています。人体には40兆個の細胞が存在しているのですよ。
主なるDNA損傷とその損傷に対する修復機構を解説しましょう。
(1)5種類の直接的なDNA損傷を修復する機構や(2)その他の5種類の間接的なDNA損傷を修復する機構説明しましょう。合計10種類のDNAの修復機構について説明しましょう。
(1)5種類の直接的なDNA損傷を修復する機構
①ミスマッチ修復。塩基対合といわれる誤った塩基対を修復。ヌクレオチド除去修復と似た方式で誤った塩基対合の周辺を広く除去して修復する。
- 塩基除去修復。軽度の塩基損傷が生じる酸化損傷、メチル化、一本鎖切断などの損傷を修復する。傷ついた塩基を含むヌクレオチドだけを除去します。
- ヌクレオチド除去修復。DNA構造を大きく変化させる損傷であり、例えば紫外線によるピリミジン二量体や化学物質による塩基付加体などを修復する。ピリミジン二量体とは、DNA鎖の隣同士のピリミジン塩基が共有結合してできる傷で、太陽紫外線(UVB)が原因で発生します。ピリミジン二量体は、皮膚がんの一種であるメラノーマの原因となる可能性が示唆されていますが、間違いです。メラノーマの原因はヘルペスによるメラニン色素細胞の増殖遺伝子の変異です。細胞にはDNA損傷を修復する機能があり、修復酵素が傷を修復します。
- 相同組み換え。DNA二本鎖切断を修復する。DNA鎖クロスリンクを修復します。DNA鎖クロスリンクとは英語でDNA-protein crosslink, 略してDPCです。正確な日本語訳はDNA-タンパク質クロスリンク (DPC)で、DNAにタンパク質が共有結合により架橋したゲノム損傷である。DNA-タンパク質クロスリンク(DPC)とは、DNAとタンパク質が共有結合によって架橋したゲノム損傷です。DNA複製や転写を阻害し、細胞に重篤な影響を与える可能性があります。DPCは、放射線や紫外線、アルデヒド化合物、重金属イオン、抗がん剤などのさまざまな因子によって誘発されます。 DPCは複製・転写装置の進行を阻害します。また修復タンパクのDNAに対する結合を阻害し細胞死を誘発することもあります。
相同組み換えとは、2本のDNA鎖の塩基配列が似ている(相同な)部位で、互いのDNA配列を交換する現象です。DNAの損傷を修復したり、遺伝的多様性を生じたりするなど、生物にとって重要な役割を果たしています。相同組換えの主な機能は①DNAの二重鎖切断(DSB)の修復②減数分裂時の染色体間の遺伝情報の交換③一部の細菌における外来DNAの取り込み④相同組換えは、原核生物から真核生物まで広く保存されており、ほぼ普遍的な生物学的機構です。⑤相同組換えの制御機構が破綻すると、遺伝情報(ゲノム)の不安定化や細胞死、ヒトにおいてはがん化の原因となる可能性があります。
- 非相同末端結合。DNA二本鎖切断やDNA蛋白クロスリンクを修復する。切断端を直接に再接合させる方式です。④の相同組み換えに比べて修復の忠実度は低いのです。修復全体に占める割合は相同組み換えよりも大きい。
(2)その他の5種類の間接的なDNA損傷を修復する機構
①DNAポリメラーゼによるDNAの損傷を正す。
染色体のDNAの複製の際に誤って取り込まれた塩基を即座に除去します。DNAの複製を行うDNAポリメラーゼはエキソヌクレアーゼ活性をもっているので間違った塩基を即座に除去できます。DNAポリメラーゼとはDNAを合成する酵素の総称です。DNAポリメラーゼは、細胞分裂の度にDNAを複製し、2つの複製品を1つずつ娘細胞に渡すことで、遺伝情報が世代から世代へと受け渡されます。エキソヌクレアーゼとは核酸(DNAまたはRNA)の末端から順にヌクレオチドを切断する酵素です。ポリヌクレオチド鎖の3’末端と5’末端のいずれかに位置するホスホジエステル結合を切断する加水分解反応を触媒します。
- テロメラーゼの機構。
染色体の末端にあるテロメアを伸長させる酵素がテロメラーゼです。短縮したテロメアやDNA二本鎖切断を修復します。癌細胞や生殖細胞でテロメラーゼの活性が高い。テロメアとは染色体の末端にあるDNAとタンパク質で構成された構造で、細胞の寿命に関与している。テロメアの役割は染色体を保護するキャップのような役割を果たし、染色体にある遺伝情報を厳重に守っています。特徴はDNAの繰り返し配列(TTAGGGなど)が約1000回繰り返された構造で、末端は一本鎖になっている。細胞分裂のたびに短くなり、年平均50塩基ずつ短くなる。生活習慣の改善により短縮のスピードを抑制できる。
テロメアが短縮すると、がん、脳卒中、心筋梗塞、糖尿病、肺疾患、認知症などの疾患リスクが高まる。
テロメアは、ギリシャ語で「末端」を意味する「テロ(telos)」と「部分」を意味する「メア(meros)」を語源としています。テロメアを伸ばす酵素として「テロメラーゼ」が存在しますが、ヒトの体細胞では発現していないか、弱い活性しか持っていません。一方、がん細胞内ではテロメラーゼが活性化されており、無限に増殖できる状態になっています。
- 細胞周期損傷チェックポイント(監視所)は3か所あります。① G1チェックポイント② G2/Mチェックポイント③ 分裂中期または紡錘体形成チェックポイント
DNA損傷に反応してDNA複製や細胞周期の進行を一時的に止めてDNA修復を促進する検問所です。
細胞周期とは細胞が増殖して分裂するたびごとの一連の繰り返しの過程、またはその周期のことを指します。細胞周期は、①G1期、②S期、③G2期、④M期の4期に分けられます。1周期で細胞は新旧入れ替えるために約24時間から50時間かかります。
- G1期。DNA合成準備期とも呼ばれ、増殖因子の有無や栄養状態などを識別して細胞分裂を進行するかを判断する。12時間~数日続く。G1期のDNA合成準備期に一番時間がかかります。
- S期。ゲノムDNAの複製が行われる。通常2~4時間続く。
- G2期。ゲノムDNAの分配が行われ、細胞質分裂の準備も行われる。2~4時間続く。
- M期。ゲノムDNAを2つの娘細胞に等分し細胞質分裂を行う。1~2時間続く。4つの厳しい間違いを正すための検問所(監視所)が細胞周期損傷チェックポイント(検問所)なのです。
細胞周期は、細胞の種類によって異なりますが、一般に増殖中の幹細胞を持っているヒトの細胞では約24時間で新旧の細胞が入れ替わります。
細胞周期チェックポイント(検問所とか監視)は、細胞が正しく細胞周期を進行させるために備わっている機構でありかつDNA修復機構の一部になっているのです。従ってゲノムDNAの遺伝子が癌化しているかどうかを精密に子細に点検しているのです。だからこそ
細胞周期チェックポイントは、細胞周期の4つの各期において、DNAの損傷や外部環境、DNA複製などの状況を監視し、ゲノムDNAの遺伝子が癌化しているかどうか異常や不具合がある場合は細胞周期進行を停止させる機構なのですが莫大な数に増えたヘルペスによるゲノムDNAの遺伝子の癌化を含むゲノムDNAの遺伝子の異常を修復でいないので、しかもDNA修復できなければアポトーシスを実行させるP53の遺伝子も異常になっても修復させることが出来ずに癌細胞が生まれてしまうのです。細胞周期チェックポイント(検問所とか監視)の働きも増えすぎたヘルペスに対しては無力で手も足も出ないのです。
我々の体の中の細胞の大部分は,組織の損傷などで細胞の再生が必要な場合を除いて,細胞分裂をしていない。しかしながら,ある限られた細胞は一定の頻度で増殖する。このような細胞を“細胞周期にある”という。細胞周期には4つの決まった順番の期が存在する。これらをG1期,S期,G2期,M期と呼ぶ。これらの器官の長さはG 1 は= DNA合成前の可変相(12時間~数日);S = DNA合成中(通常2~4時間);G 2 = DNA合成後(2~4時間)―細胞内にDNAの四倍体がみられる;M 1 = 有糸分裂(1~2時間)。細胞周期の進行は3つのチェックポイント(check points)で監視されている。
下図に3つのチェックポイント(check points)である① G1チェックポイント② G2/Mチェックポイント③ 分裂中期または紡錘体形成チェックポイントと、色分けされた4つの細胞周期であるG1期、S期、G2期、M期を示しました。G1期から2重点線で書かれたG0期とは休止期と呼ばれ細胞増殖なしで増殖能力は保ちつつも細胞分裂を停止している状態のことです。
1個の細胞について3つのチェックポイント(check points)と1個の細胞が2個の娘細胞に増殖して分裂するまでの4つの細胞周期であるG1期、S期、G2期、M期のプロセスを数に示します。

① G1チェックポイント:栄養や増殖因子が存在するか?DNAの修復は完了したか?
② G2/Mチェックポイント: 染色体DNAが分配が可能か? DNA複製は完了したか? DNA損傷の修復は完了したか?
③ M期の分裂中期または紡錘体形成チェックポイント:紡錘体の形成は完了したか?
紡錘体とは細胞分裂の際に染色体を娘細胞に分配するために形成される細胞骨格構造です。細胞分裂装置とも呼ばれます。
これらのチェックポイント(検問所)では2種のタンパク質 であるサイクリン(cyclin)とサイクリン依存性キナーゼ(cyclin-dependent kinase略してCDK訳してサイクリンリン酸化酵素)から成る複合体が中心的役割を果たすのです。左にM期に行われる染色体の「有糸分裂」を前期(prophase)→中期(metaphase)→後期(anaphase)→終期(telophase)までの染色体が2つの娘細胞に分配される様子を示しました。同時に紡錘体と分裂中期と分裂後期の紡錘体形成チェックポイントの様子も示しました。上の図のG0とは何でしょうか?G0期(ジーゼロ期)とは、細胞が栄養状況が悪化したり環境が増殖に適さなくなったりしたときに、増殖能力を保ちつつも細胞分裂を停止する期間です。しかしG0期の細胞は、通常の機能を果たし続けが、細胞分裂を一時的または永久的に停止することがある。神経細胞や筋細胞などの分化した細胞は通常G0期にある。増殖刺激が与えられると、S期を経て細胞周期の進行を再開する。G0期には、可逆的な状態(静止状態)と不可逆的な状態(老化状態と分化状態)の3つの状態が存在します。G0期の長さは12時間から3日間です。
このようにDNA損傷の修復のために細胞周期チェックポイントでDNA損傷があるかどうかを監視していれば「サイクリン-サイクリン依存性キナーゼ複合体」の働きを使ってヘルペスウイルスによるDNAの損傷以外のすべてのDNAの損傷を修復してしまうのです。
何故ヘルペスによるゲノムDNAの損傷(変異)、つまりゲノムDNAの蛋白質の設計図である遺伝子の損傷(変異)だけは修復できないのでしょうか?人体は40兆の多細胞から成り立っています。上に掲載した(体)細胞周期の図は40兆の細胞の1個の細胞についての説明図であります。1個の細胞には父と母の各々1本の染色体が一対になった2倍体(一対の2本の染色体)の相同染色体が23本存在しています。1倍体(一本の染色体)で染色体
を数えると46本の相同染色体になります。相同染色体とは生物の体細胞の核内に存在する、大きさや形状が同じで、塩基配列もほぼ同一の2本の染色体で、一方の染色体は母方から、もう一方の染色体は父方から受け継いだものです。ところが全く同じ細胞を増やすために分裂してすべてが同じ部品でできているなにもかも遺伝子もまったく同じ細胞を2倍にする必要があります。まずは46本の遺伝子が乗っている染色体を2倍に増やす必要があります。そのためにバラバラになった46本の染色体をコピーして2倍のバラバラになった92本の姉妹染色分体にします。
バラバラになる前の染色体(クロモソーム)の中ではDNAはクロマチンという形で存在しその基本単位がヌクレオソームなのです。ヌクレオソームの役割はDNAを安定に細胞核内に収納する。DNAをヒストンから解離したりヌクレオソーム同士の集合状態を変えたりすることで、遺伝子発現のON/OFFスイッチを調節する。遺伝子の活動を部分的に変更する。ヌクレオソームとは細胞内のDNAを保護しながら、DNAの情報を利用できるようにする役割を担う構造で、ヌクレオソームは、中央に8つのヒストンタンパク質が束ねられ、その周囲を2つの環状のDNAが取り囲む構造をしています。このヌクレオソームのがっちりした構造により、DNAをぎっしり詰め込んで損傷しないようにする強固な保護的構造が作られます。
遺伝子は染色体の上に決まった順序で並んでいます。人の場合には1個の細胞には46本の染色体に約23000個の遺伝子が振り分けられているので、染色体1本あたりの遺伝子数は約500~2800個の範囲になります。
つまり対になっていた染色体は元の1個の細胞の染色体46本×2で92本の姉妹染色分体ができます。増殖分裂の細胞周期の4期の順序の概略であり1個の体細胞の染色体は46本ありますがの分裂の経過についてはM期だけの詳細以外は他の3つの細胞周期のG1期とS期とG2期の染色体が分裂して何倍体の染色体になったのかについては一切触れられてはいません。
HerpesがゲノムDNAに感染する機会はG1期の父親と母親のそれぞれ1本の相同染色体だったのが複製されて同じ染色体が2倍になって姉妹染色分体に増えます。
上の図でG1期から2重点線で書かれたG0期とは何でしょうか?G0期とは増殖能力は保ちつつも細胞分裂を停止している状態のことを指します。 実は、体内に存在する細胞のほとんどはG0期にとどまっており、増殖を停止しています。 これは細胞が接触しているために生じる性質として考えられておりコンタクトインヒビション(細胞の接触障害)と呼ばれています。G0 期は 12 時間から3日間続く。
細胞周期チェックポイントの検問の例としては、①G1/Sチェックポイント:栄養や増殖因子が存在するか、DNAの修復は完了したかなどを確認します。②G2/Mチェックポイント:染色体DNAが分配が可能か、DNA複製は完了したか、DNA損傷の修復は完了したかなどを確認します。
分裂中期または紡錘体形成チェックポイント:紡錘体の形成は完了したかなどを確認します。
細胞周期チェックポイントの機能は、正確な遺伝情報を娘細胞、ひいては子孫に伝達するために重要です。この機構の異常はヒトなどのがん発生の主要な原因のひとつといわれています。
Cre-loxP部位特異的組換えは別のもう一つの組み換えでありバクテリオファージ(細菌ウイルス)が宿主のDNAに入り込むときに使う機構です。Cre-loxP部位特異的組換えは、1981年にバクテリオファージ(細菌ウイルス)のP1の研究で見出された部位特異的組換え反応である。loxPという特定のDNA配列を標的としており、DNA組換え酵素Creにより触媒される。条件的遺伝子ノックアウトを実施する目的などで広く使われる技術となっている。
Cre-loxP部位特異的組換えの原理は何か?バクテリオファージP1が大腸菌内で増殖する際には、ファージ(細菌ウイルス)のゲノムを環状化し複製するためにこの組換えシステムを利用している。この反応に必要な構成要素は組換え酵素のCreと標的配列のloxPだけである。
組換え酵素Creとは何か?creはcauses recombinationの英語の略で「組み換えをおこす酵素」という意味です。Creタンパク質は部位特異的組換え酵素であり、DNA分子の特定の配列(loxP)同士の間で組換えを起こす。343残基からなり分子量はおよそ38 kDaである。 チロシンリコンビナ)ゼファミリーに属しており、アミノ末端側の一部分を除けばラムダファージのインテグラーゼと良く似た構造をしている。creはcauses recombinationの英語の略です。なお、タンパク質名がCreであり、遺伝子名は小文字のcreである。
反応機構はおよそ次のとおりである。まずCreの単量体2つがloxP配列両端の13 bp(
13対の塩基)配列をそれぞれ認識し、それが2組集合することで反応の場が形成される。Creの324番目のチロシンがloxPの切断部位を攻撃して鎖交換を触媒し、ホリデイ・ジャンクションの異性化を経て、再び同様の鎖交換が行われる。ホリデイジャンクション( Holliday junction)とは、4つの核酸配列が塩基対形成によって保持されている核酸構造である。
標的配列loxPとは何か?loxはlocus of crossing overで遺伝子を交差するという意味です。に、Pはファージ(phageのP1)に由来している。
loxPは元来バクテリオファージP1のゲノム中にある34塩基からなる配列であり、 中央8 bpは非対称な配列でその両側13 bpが対称になっている。loxはlocus of crossing overに、PはファージP1に由来している。
Cre結合部位 組換え部位 Cre結合部位
ATAACTTCGTATA – G^CATACAT -TATACGAAGTTAT
応用技術について。
このCre-loxP部位特異的組換え技術によって、研究者は研究対象の生物の遺伝子発現を制御したり、特定のDNA配列を除去したり、染色体構造を改変したりすることが容易になった。
Creは一般には存在しない酵素であるため、遺伝子組換え技術により利用したい細胞でCreが生産されるようにする必要がある。言い換えれば、発現されるように細工した細胞以外では組換えが起こらない。またloxP配列も34 bpと長いため、通常は内在していないと考えられる。したがって、特定の場所にloxP配列を用意すれば、その場所以外で組換えが起きる可能性は極めて低い。

細胞がCreを生産すると、その細胞のゲノム中にあるloxP部位同士の間で組換えが起きる。 1つのDNA分子に2つのloxP配列がある場合、その向きによって結果が変わる。 loxP同士の向きが逆であれば挟まれた配列は反転するが、同じ向きだと挟まれた配列が切り出される。 これによって例えば染色体の一部を転置・欠失させたり、染色体同士を転位させたりすることができる。

loxP配列が逆向きの場合 loxP配列が同じ向きの場合
条件的ノックアウト
個体の発生に必須な遺伝子は単純な方法ではノックアウト個体を得ることができないが、Cre-loxP系を用いることで特定の条件下でのみ遺伝子ノックアウトすることが可能になる。
発現特性が既知のプロモーターを利用して、Creを特定の組織あるいは特定の時期に生産するマウスを用意する。 例えばもし脳でだけOFFにしたいなら、そこでだけ機能するプロモーターをさがし、 その後ろにCreの遺伝子creを繋げて、脳でだけCreが生産されるようにする。 次に、目的遺伝子の前後にloxPを挿入したマウスを用意する。 この2つのマウスを交配させて得られた子孫のうち、双方の遺伝子を持つものを選択する。
Cre-loxPシステムによるノックアウトマウス作成の概略。creを遺伝子導入したマウス(左上)と、解析対象の遺伝子をloxPで囲み下流にGFP遺伝子を配置したマウス(右上)とを掛け合わせる。creが発現した細胞(左下)では解析対象の遺伝子が排除されるため、仮にその遺伝子が発現すべき時には代わりにGFPが発現する。creが発現しない細胞(右下)では元々の遺伝子が発現する。どの細胞でいつcreが発現するかはcreのプロモーターが決定する。
使いたいプロモーターが胚発生の初期にも活性化してしまうことはよくある。 そんな場合のために、creのプロモーターを改変してドキシサイクリンやタモキシフェンなどの薬剤を使って活性化を誘導出来るようにすることで、確実にノックアウトを制御できる系がある。
条件的遺伝子修復
loxPの向きを調整することで、遺伝子を排除してしまうのではなく、 逆位させて機能を止めておき、必要に応じて再び逆位させて機能を回復させることもできる。
選択マーカーの除去
遺伝子改変をする際に、薬剤耐性などの選択マーカーを利用することがよくある。 しかしこうした選択マーカーの存在が実験結果をゆがめる可能性が存在する。 そこで、マーカー遺伝子の前後にloxPを配置して遺伝子改変を行い、 目的の改変個体を得た後でcreの発現を誘導すれば、選択マーカーを除去することができる。
3. PKM1による腫瘍促進作用の発見
筆者らのグループは,前節の問題を解決し,PKM1およびPKM2のアイソフォーム特異的な機能を明確にするため,新たなPKM遺伝子改変マウスであるPKMノックインマウスを開発した。これら変異マウスでは,内在性PKM遺伝子座に,PKM1あるいはPKM2をコードするcDNAがノックイン(KI)挿入してある(図6).そのため,これら変異アリルからは,PKM1(PkmM1から)あるいはPKM2(PkmM2から)の,いずれか片方のアイソフォームしか発現しない.重要なのは,それら変異アリルに由来する各アイソフォームの発現は,基本的にそろっているという点である.
Cre/loxPシステムをもっと詳しく勉強し完全にマスターしましょう。
Cre/loxPシステムとはloxP配列と呼ばれるDNA配列に対しDNA組換え酵素Creが働くことにより生じる部位特異的組換え反応を利用した遺伝子組換え実験系である。本来、バクテリオファージP1が宿主である大腸菌内で複製される際に自身のゲノムを環状化するための組換えシステムである。遺伝子工学、発生工学に分野で主にloxP配列間に存在する遺伝子をCre発現により欠失させるために使用される。
Cre/loxPシステムとは
Cre/loxPシステムとはloxP配列と呼ばれるDNA配列に対しDNA組換え酵素Creが働くことにより生じる部位特異的組換え反応を利用した遺伝子組換え実験系である。この組換え反応は主に特定のDNA配列の除去に用いられるが、現在ではこの技術を応用して後述のCre発現依存的遺伝子欠損やCre発現依存的遺伝子発現など遺伝子発現実験などにも利用されている。
基本要素

図1.loxP配列が同方向の場合のCreによる組換え

図2.loxP配列が異なる方向の場合のCreによる組換え
Creレコンビナーゼ
バクテリオファージP1に由来し、TypeIトポイソメラーゼに分類される約38kDaのDNA組換え酵素でloxP配列とよばれる塩基配列を認識し組換えを起こす。バクテリオファージP1のゲノムは、宿主である大腸菌内で複製される際に環状化するために組換えシステムを利用しています。この組換えシステムはCre/loxPシステムと呼ばれ、遺伝子工学や発生工学の分野で利用されています。バクテリオファージは細菌に感染して増殖するウイルスの総称で、T4ファージなどが有名です。T4ファージとは
loxP配列
バクテリオファージP1ゲノムに由来する34bpのDNA配列で両末端に存在するCre結合部位13bpは対称に、中心部は非対称な配列になっている。
5′-ATAACTTCGTATAGCATACATTATACGAAGTTAT-3′
CreとloxP配列の反応
2つのloxP配列が同方向に位置している場合、CreによりloxP配列間の遺伝子は切り出され環状化する(図1)。この反応は平衡反応であるが切り出され環状化した遺伝子断片が元の状態に戻ることは極めて稀である。
また、2つのloxP配列が異なる方向に位置しているときはCreによりloxP間の遺伝子は反転する(図2)。
バリエーション
Creのバリエーション
Creと変異エストロゲン受容体の融合タンパク質であるCre-ERタンパク質は通常細胞質に存在するが、エストロゲン誘導体であるタモキシフェンと結合することにより核内に移行し、loxP配列に対して組換えを起こす。これを利用してCre-loxPシステムの働く時期をタモキシフェン依存的に調節することが可能である(図3)。

図3.タモキシフェンによるCre-ERの制御
loxP配列のバリエーション
Creによって組換わるものの、loxP配列とは組換わらないlox511, lox2272, loxFASなど種々の変異lox配列
lox511: 5′-ATAACTTCGTATAGtATACATTATACGAAGTTAT-3′
lox2272: 5′-ATAACTTCGTATAGgATACtTTATACGAAGTTAT-3′
loxFAS: 5′-ATAACTTCGTATAtacctttcTATACGAAGTTAT-3′
(小文字はloxP配列と異なる塩基)
や染色体への遺伝子挿入が可能なlox RE, loxLE配列
lox RE: 5′-ATAACTTCGTATAGCATACATTATACGAAcggta-3′
lox LE: 5′-taccgTTCGTATAGCATACATTATACGAAGTTAT-3′
(小文字はloxP配列と異なる塩基)
などが存在する。
Cre/loxPと類似のシステム
出芽酵母(Saccharomyces cerevisiae)由来の組換え酵素FLPとFRT配列
5′-GAAGTTCCTATTCTCTAGAAAGTATAGGAACTTC-3′
もCre-loxPシステムと同様の目的で頻用される。 また、醤油酵母(Zygosaccharomyces rouxii)由来の組換え酵素RとRS配列、バクテリオファージMu由来の組換え酵素Ginとgix配列も例は少ないものの使用されている。
使用例
Cre発現依存的遺伝子欠損(コンディショナルノックアウト)
特定の遺伝子の機能をマウスなど動物個体で解析することを目的として、その遺伝子を欠損させた遺伝子ノックアウト動物が用いられる。しかし対象とする遺伝子の欠損により致死となるため目的の解析が出来ないことがある。そのような場合はある特定の組織だけで遺伝子を欠損することでこの問題を回避できる。 まず、欠損させたい遺伝子の両端にloxP配列をジーンターゲティング法により挿入した遺伝子改変動物を作製し、さらに組織特異的プロモーターの制御下でCreを発現するトランスジェニック動物を交配する。得られた動物個体内では組織特異的プロモーターが働く部位のみ遺伝子が欠損する。また、Creを発現するためのプロモーターを選択することにより、組織特異的な遺伝子ノックアウトだけではなく時期特異的な遺伝子ノックアウトも可能である(図4)。

図4.Cre発現依存的遺伝子欠損
Cre発現依存的遺伝子発現
ある遺伝子を細胞あるいは動物個体の組織で高発現させるためには組織特異的プロモーターの下流に目的の遺伝子をつなげた外来遺伝子を構築し導入する方法が用いられる。 しかし、そのプロモーターによる発現量が低かった場合には以下の方法を用いることでプロモーターの組織特性を保ちながら目的遺伝子の高発現が期待できる。 まず、組織特異的プロモーターの下流にCre遺伝子をつけた外来遺伝子およびサイトメガロウイルス(CMV)プロモーターなどの高発現が期待できるプロモーター下流に2つのloxP配列に挟まれたpolyA付加シグナルなどの転写停止配列、さらにその下流に目的の遺伝子をつなげた外来遺伝子をそれぞれ構築する。これらの外来遺伝子を同時に導入すると、組織特異的プロモーターが働く部位ではloxP配列間の転写停止配列が欠損し、その下流に位置する目的遺伝子が高発現する。一方、組織特異的プロモーターが働かない部位では転写停止配列が存在するため目的の遺伝子は発現しない(図5)。

図5.Cre発現依存的遺伝子発現
培養細胞における選択マーカーの除去
培養細胞に外来遺伝子を導入する際、細胞の遺伝子が導入された細胞を選択をするためにネオマイシン耐性遺伝子など薬剤耐性遺伝子を選択マーカーとして用いる。しかし、目的の細胞が得られた後にはこれらの選択マーカー遺伝子を発現するためのプロモーターが内在性のプロモーターに干渉する例があるために、選択後は除くことが望ましい。この場合にもCre/loxPシステムが利用される。細胞に導入する外来遺伝子の選択マーカーは2つのloxP配列で挟まれた形にしておく。この選択マーカーを除くためにCreとともにピューロマイシン耐性遺伝子を発現するプラスミドpCrePacを一過性に導入し、ピューロマイシン存在下で培養するとCreにより選択マーカーが除去された細胞のみが生存する(図6)。

図6.培養細胞における選択マーカーの除去
3. PKM1による腫瘍促進作用の発見
前節の問題を解決し,PKM1およびPKM2のアイソフォーム特異的な機能を明確にするため,新たなPKM遺伝子改変マウスであるPKMノックインマウスを開発した。これら変異マウスでは,内在性PKM遺伝子座に,PKM1あるいはPKM2をコードするcDNAがノックイン(KI)挿入してある(図6).そのため,これら変異アリルからは,PKM1(PkmM1から)あるいはPKM2(PkmM2から)の,いずれか片方のアイソフォームしか発現しない.重要なのは,それら変異アリルに由来する各アイソフォームの発現は,基本的にそろっているという点である.アレル変異とは何ですか?同一遺伝子座に属し、互いに区別される遺伝的変異体をアレル変異と呼びます。

図6 Pkmノックインの模式図。ノックインとは、外来遺伝子を標的とするゲノム領域へ挿入することを指します。ノックアウトとは、特定の遺伝子を破壊することを指します。
ノックインとノックアウトは、遺伝子改変マウスなどでも使われます
上記変異マウスの発がんポテンシャルを調べるため,Pkm-KIに加えKrasLSL-G12D変異を併せ持つPkm;KrasダブルKIマウスを作製した.これらマウスにCre発現アデノウイルスを気管内投与し,肺がんを誘導した.驚いたことに,Pkm1-KIマウスでは,Pkm2-KIや野生型マウスと比べ,肺がん形成が促進されていた.Pkm-KIの効果をより広範に調べるため,新生仔マウスに発がんイニシエーターであるDMBAを塗布し,経過を観察した.通常,野生型マウスにDMBAを単独で塗布しても,腫瘍形成まで至ることはない.一般的には,DMBAに加えて何らかの発がんプロモーター(TPAやオカダ酸など)が追加的に必要とされる.ところが,Pkm1-KIマウスでは,肝がんを中心に,さまざまな腫瘍性病変をみとめた.野生型マウスやPkm2-KIマウスでは,そのような病変は一切みとめられなかった.これら結果は,Pkm1に発がんプロモーターとしての作用があることを強く示している.一連のマウス動物実験により,Pkm1が,Pkm2よりも,強いがん促進作用を持つことが示された.
Pkm1によるがん促進作用が,腫瘍細胞自律的なものか否かを調べるため,Pkm-KIマウスからマウス胎仔線維芽細胞(MEF)および肺上皮(LE)細胞を単離して不死化したのち,発がん変異型KrasやEGFRで形質転換し,解析を行った12).培養系においては,形質転換MEFおよびLE細胞の増殖に,Pkmの遺伝子型による差異はみとめられなかった.ところが,これら細胞をヌードマウスに移植し,その腫瘍増大を比較したところ,Pkm1-KI腫瘍では,Pkm2-KIおよび野生型腫瘍に比べ,腫瘍増大が早かった.これら実験により,Pkm1が,腫瘍細胞自律的に,腫瘍増殖を促進できることが示された.
4. PKM1によるグルコース代謝亢進とワールブルグ効果解消
PKM1による腫瘍促進の分子機構を探るため,Pkm1およびPkm2-KI細胞の代謝を比較することにした.13C標識グルコースを用いたトレーサー実験を行ったところ,PKM1が,グルコース由来炭素のTCA回路への流入および乳酸への変換,そのいずれもを促進させることがわかった.重要なことに,Luntらが提唱したモデルとは異なり,PKM1によって核酸や各種アミノ酸の生合成が妨げられることはなかった。
このとき,もう一つ,興味深い観察結果が得られた.それは,PKM1はPKM2と比べ,グルコースからの乳酸産生を上昇させる一方,総乳酸産生を減少させるというものであった.この結果は,Christofkらの報告にきわめて類似しており,あらためて,PKM2がワールブルグ効果様の性質に寄与していることを再確認することができた.同時に,PKM2発現細胞でみられる乳酸産生の増加は,グルコース以外の炭素源に由来している可能性を示していた.
そこで,13C標識グルタミンを用いたトレーサー実験を行った結果,Pkm2-KI細胞では,Pkm1-KIと比べ,グルタミンが,より活発に,アスパラギン酸やピルビン酸・アラニン・乳酸へと代謝されていることがわかった.これら結果と一致して,PKM2が,グルタミン消費を亢進させる結果も得られた.
一連の代謝解析によって,PKM1が,各種生体成分の生合成を妨げることなくグルコースの異化を亢進すること,一方で,栄養源としてのグルタミン利用を減弱させることが明らかになった.そのような代謝ネットワークのリプログラミングが,未解明の制御メカニズムを通じて,結果的に総乳酸産生を低下させていると思われる.
5. 小細胞肺がんのPKM1依存の発見
我々がPkm-KIモデルを使って得た一連の検討結果は,もし腫瘍細胞に発現すれば,PKM1の方が,PKM2よりも,強いがん促進作用を持つことを示していた.しかし,これまでのところ,PKM1を発現するヒトがんは知られていなかった.我々はこれを,ほとんどのがん起源が元来PKM2発現型だからではないか?と考え,PKM1を発現する正常細胞に着目した.そのうちの一つが,気管支の神経内分泌細胞である.重要なことに,この細胞は,肺がんのなかでも高悪性で知られる小細胞肺がん(small-cell lung cancer:SCLC)の起源と考えられている.SCLCは,全肺がんの15~20%を占めている.他のタイプのがんと比べ,初回の抗がん剤治療・放射線治療が効きやすい傾向にあるが,その大半が再発を来し,予後はむしろ悪い.大半の症例でTP53とRB1遺伝子の同時機能喪失がみとめられているが,これまでに創薬可能なドライバー遺伝子異常は発見されていない.
手術検体および細胞株を用いた検討などにより,SCLC(小細胞肺がんsmall-cell lung cancer:SCLC)では,PKM1発現が高く,PKM1/PKM2比も,他がん種と比べて圧倒的に高いことがわかった.質量分析にもとづいた絶対定量により,タンパク質レベルでのPKM1/PKM2比を肺がん細胞株間で比較したところ,SCLCでの値は,非SCLCのそれと比べて10~20倍程度も高いことがわかった.
SCLCにおけるPKM1発現の意義を調べるため,ヒトSCLC細胞株を親株として,PKM1あるいはPKM2のどちから一方のみ発現する細胞株を作製し,増殖を調べた.その結果,PKM2のみを発現する細胞では,PKM1のみを発現する細胞と比べ,増殖が著しく減弱されていた.つまり,PKM2では,PKM1の役割を相補できないことがわかった.これら結果から,少なくとも検討したSCLC株においては,PKM1に起因する高いPKM活性が,増殖・生存に必須であることが示唆された.
6. まとめ
Pkm-KIマウスを使った今回の検討により,PKM1は,PKM2よりも,1)各種生合成経路を減弱させることなく,グルコース代謝を亢進させ,2)腫瘍増殖を促進させることができることがわかった.これら結果は,既存の“PKM2によるグルコース異化の制限が、がんに必須である”という定説を覆すもので,ワールブルグ効果の役割についての再考が必要となるだろう.ワールブルグ効果は,腫瘍細胞自律的には,がん細胞の増殖にむしろ不利に働くことを示唆している.よって,もしワールブルグ効果に腫瘍促進的な側面があるとすれば,それはヘルペスウイルスによる非細胞自律的なものではないかと考えている.がん代謝と微小環境との相互作用などについて,今後,さらなる解析が必要と考えている.
これらの問題とは別に,SCLCがPKM活性の低下という代謝干渉に対して脆弱であるという点は,大へん興味深いと思っている.SCLCの治療をめぐっては,この15~20年の間,根本的な進歩はほとんどなかった.この難治性肺がんの場合,現時点で創薬可能なゲノム変異がほとんど存在しないことも明らかになっており,非ゲノム性の創薬アプローチの重要性が指摘されていた.今回,PKM1やその関連形質が,SCLCの新たな治療標的となる可能性が示された.現在,それらを標的とする新規治療の開発に取り組んでいる。
代謝干渉とは代謝妨害するという意味合いが含まれている新しい概念です。代謝とは、体内で酸素や栄養素を取り込み、消化・吸収してエネルギーや活動に必要な物質を産生する化学反応です。また、体内の活動で不要になった老廃物を排出する働きもあります。
代謝には、次のような種類があります。①基礎代謝: 生命を維持するために必要なエネルギーで、呼吸や心臓の動きなどに使われます。基礎代謝が高いと、血行が促進され、脂肪燃焼が活発になる効果が期待できます。②活動代謝: 体を動かすことによって消費されるエネルギーです。③食事誘発性熱生産: 食事したものを消化するために使われるエネルギーです。
代謝が下がる原因としては、①不規則な生活(暴飲暴食やストレスなど)②過度な食事制限(極端なダイエットや偏った食事など)
部位特異的組み換えはλファージが大腸菌DNAでも行われているので簡単に説明しましょう。又λファージはヘルペスウイルスと同じように溶原サイクルと溶菌サイクルをもっています。
λファージ(ラムダファージ)が大腸菌のDNAと相互作用する様子は、次のとおりです。
大腸菌に感染すると、λファージの直鎖2重鎖DNAの両端にあるcohesive endが水素結合して環状になります。
その後、λファージのDNAは溶菌サイクルまたは溶原サイクルのいずれか一方のサイクルに入ります。
溶原サイクルでは、λファージ(ラムダファージ)のDNAが大腸菌の染色体に組み込まれ、大腸菌DNAの一部として複製されます。これは人の細胞でヘルペスもやっていることで人の染色体に組み込まれてDNAの一部となって複製されてしまうとDNAの遺伝子が癌遺伝子になったりするのです。
λファージの溶菌サイクルでは、溶菌サイクルのλファージ(ラムダファージ)は、大腸菌に感染した後に宿主の複製・発現機構を利用してファージDNAを複製し、外殻タンパク質を合成してラムダファージ粒子を形成して増えていくウイルスでヘルペスウイルスにそっくりです。しかも最後は大腸菌の単細胞を殺してしまうのも人の細胞に感染したヘルペスウイルスとそっくりです。ラムダファージ粒子が進化してヘルペスウイルスが誕生したかのようです。
λファージの溶菌サイクルの主な流れは次のとおりです。大腸菌の染色体DNAに組み込まれていたプラスミドDNAが切り離される。プラスミドDNA とはプラスミドDNA(pDNA)とは、染色体とは独立して存在し、独立して複製する小さなDNA分子です。大腸菌などの細菌や酵母の核外にでて存在し、一般的には環状の2本鎖構造をしています。プラスミドDNAは、次のような特徴があります。①細胞分裂によって娘細胞へ引き継がれるのはヘルペスと似ています。②薬剤に対する耐性を示すタンパク質の遺伝子を持つものや、③細菌の接合を起こすものなど、さまざまな役割を持つ。④細菌が特殊な環境に置かれた場合や病原性を発揮する場合等に、プラスミドの遺伝子が独自に働く。herpesは細胞分裂や免疫が低下したときに遺伝子が独自に働いて増殖するのも似ています。遺伝子組み換え操作のベクターとして利用される。プラスミドとは、細菌や酵母の細胞核外に存在するDNA分子の総称で、染色体DNAとは独立して複製する能力を持っています。環状または線状の2本鎖構造をしており、細胞分裂によって娘細胞へ引き継がれます。このような特徴はすべてヘルペスが持っています。ベクターとは遺伝子組み換え操作で遺伝子を運ぶ媒体でラテン語の「運び屋(vehere)」に由来し、ヒト細胞や核に目的遺伝子を組み入れるために使用されますが、Herpesは自分勝手にヒト細胞や核に目的遺伝子を組み入れ遺伝子を勝手気ままに組み替えています。この意味ではヘルペスは同時にウイルスベクターやプラスミドベクターの役割を自分のために感染した細胞や核や染色体でやりたい放題行っているのです。研究現場では最終的な目的に合わせてベクターを選択されますがヘルペスは自由自在に自分のをせ子供であるビリオンを作ってたのしみ最後は人を殺すことまでやっているのです。
λファージは宿主の複製・発現機構を利用してファージDNAを複製し、外殻タンパク質を合成したりするのもヘルペスがエンベロープの外殻タンパク質を合成するのと全く同じなのでそっくりです。ファージ粒子(ヘルペスのビリオン粒子と同じ)を形成し、十分な数が出来上がるとリゾチームを合成する。リゾチームによって大腸菌の細胞壁が破壊され、ファージが外に飛び出して大腸菌を殺してしまいます。λファージはherpesの祖先といってもいいのです。
λファージは、大腸菌に感染すると、ウイルス粒子内の直鎖2重鎖DNAが環状になります。herpesも環状になることもあります。その後、溶菌サイクルと溶原サイクルのどちらか一方のサイクルに入ります。
ラムダファージは、細菌や古細菌に感染して複製するウイルスで、正式にはバクテリオファージと呼ばれます。ファージは「食べる」という意味で、バクテリアを食べることから名付けられました。herpesも癌細胞や正常な細胞に必要な栄養素のすべてを奪い取りいわば細胞を食べつくしてヘルペスだけが無限大に増えてヘルペスだけが生き残り反映するのはまさに細胞ファージというべきなのです。
DNAが同方向に連結した直鎖状多量体とも呼ばれるconcatamerとなりファージの殻の中に詰め込まれます。concatamer とは、ラムダファージなどの短いゲノムDNAが複数直鎖状に連結したもので、直鎖状多量体といいます。ファージなどの短いゲノムDNAがいつくか直鎖状に連結したものです。ラムダファージが作ったたくさんの短い子供のゲノムDNAが
集合して生まれたのがconcatamerなのです。バクテリアにおける遺伝子破壊株の作製は、ファージのDNAが大腸菌の菌体内で合成されるのです。λファージのゲノムサイズは48,502塩基対ですがherpesのゲノムサイズは約150000塩基対です。
T4ファージについても説明しましょう。
腸内細菌ウイルスT4 は大腸菌に感染するバクテリオファージの種である。二本鎖DNAウイルスに分類され、ミオウイルス科に属する。T4ファージの感染は必ず溶菌を起こし、溶原化しない。過去には他のファージ種や系統であるT2ファージやT6ファージと合わせて、T偶数ファージとも呼ばれていた。
腸内細菌ウイルスT4 (T4ファージ)とは大腸菌に感染するバクテリオファージの1種である。二本鎖DNAウイルスに分類され、ミオウイルス科に属する。T4ファージの感染は必ず溶菌を起こし、溶原化しない。過去には他のファージ種や系統であるT2ファージやT6ファージと合わせて、T偶数ファージとも呼ばれていた。
バクテリオファージの語源は「細菌を食べるもの」を意味し、ファージは偏性細胞内寄生体で宿主の細胞内で増殖し、溶菌とともに細胞外へ放たれる。160種ほどの遺伝子を持つT4ファージは、ウイルスとしては最大級の大きさを持ち、モデル生物として重用されてきた。現在に至るまでT4ファージはウイルス学と分子生物学の発展に重要な役割を果たしてきている。
T4ファージウイルス粒子の構造はT4ファージはウイルスとしては大きく、ほとんどのウイルスの長さが25nmから200nmの長さであるのに対し、幅がおよそ90nm、長さが200nmである。DNAのゲノムはカプシドと呼ばれる二十面体の頭部に格納される。尾部は空洞であり、細菌の細胞に吸着したファージが、細胞内に核酸を送る際に核酸が尾部を通過する。尾部は宿主の細胞表面に存在する受容体の認識において重要であり、吸着した細菌が宿主域の範囲であるか否かを決定する。
T4ファージの尾部の末端である6MDaの基盤は、13種類のタンパク質(遺伝子産物5、5.4、6、7、8、9、10、11、12、25、27、48、53)の組み合わせからなる、全127本のポリペプチド鎖で構成される。近年原子レベルで詳細な構造が明らかにされた。尾管の近位領域はgp54が構成しているが、主な部分はgp19が構成している。ものさしタンパク質gp29は基盤-尾管複合体に存在するが、モデル化されていない。
T4ファージの大腸菌感染の過程はT4ファージはlong tail fiber (LTF) が大腸菌の細胞表面に存在するOmpCポリンタンパク質とリポ多糖 (LPS) に結合することで感染する。認識シグナルはLTFを通じて基盤に送られ、これによりshort tail fiber (STF) が不可逆的に大腸菌の細胞表面に結合する。基盤の構造変化と筒状の構造物であるtail sheathの収縮により、尾管の末端にあるGP5が細菌の細胞外膜に穴を開ける。GP5のリゾチームドメインが活性化し、細胞表層のペプチドグリカン層を分解する。残りの膜成分も分解を受けると、頭部に格納されたDNAが尾管を通って大腸菌の細胞内へ侵入する。
T4ファージの増殖はファージが細菌内に侵入して細胞を破壊するまでの溶菌サイクルは37℃でおおよそ30分程度で終了する。ビルレントファージは細菌宿主に感染すると直ちに自己増殖を開始する。ビルレントファージとは特定の細菌に感染して増殖し、その細菌を溶かす溶菌活性を持つバクテリオファージ(細菌ウイルス)の一種です。 T4ファージの子ウイルスの数が一定の量に達すると、T4ファージは宿主を溶解して破壊し、菌体外へ放出されて次の宿主細胞へと感染する。この宿主の溶解とファージの放出を溶菌サイクルと呼ぶ。この溶菌サイクルもヘルペスウイルスと同じです。溶菌サイクルとは感染した細胞とその細胞膜の破壊を伴うウイルスの増殖過程なのです。そのため、ウイルスは増殖と宿主細胞への感染のために以下の5つの過程を行います。T4ファージはラムダファージにも非常に似ています。ということはT4ファージはヘルペスウイルスにも似ています。
- 最近に吸着して侵入します。②宿主の遺伝子発現をさせない。③酵素の合成を行います。④DNAの複製を行います。⑤新しいファージウイルス粒子を産生するというの5つの過程があります。Herpesウイルスとそっくりです。
新しいファージの粒子の合成が完了すると宿主の細菌は破壊され新生ファージを環境中に放ち、宿主の細菌細胞は崩壊に至る。菌体が崩壊した際に放出される子孫ウイルスの数をburst sizeと呼び、T4ファージの場合は感染した1個の大腸菌体当たりのburst sizeは100-150個である。herpesウイルスの一個の細胞当たりのburst sizeはおよそ数百から1000個といわれます。
T4ファージの吸着と侵入のDNA注入の過程は他のファージと同様に、T偶数ファージはランダムに宿主の菌体表面に吸着するわけではない。T偶数ファージとは大腸菌に感染するT2、T4、T6などの偶数番号のファージを指します。これらのファージは、性質や形態、DNA分子の多くの部分に類似点があり、タンパク質の外殻をもち、頭部にはDNAを保持しています。大腸菌に感染すると、DNAのみを菌体内に注入し、大腸菌のDNAを破壊して子ファージを増殖させます。
一方、とは、大腸菌に感染する病原ファージの総称で、T1からT7までの7種類があります。T1、T3、T5、T7などの奇数番号のファージはT奇数ファージと呼ばれ、T偶数ファージと比べてあまり類似性がありません。
T系ファージは菌体の表面にある、特定の構造を持つタンパク質の受容体に選択的に結合する。受容体の種類はファージの種ごとに異なり、タイコ酸、細胞壁を構成するタンパク質やリポ多糖、鞭毛、線毛など多岐にわたる構造が受容体としてファージとの結合に寄与する点は全ての細胞膜に結合するヘルペスウイルスとは異なります。タイコ酸とは、グラム陽性細菌の細胞壁に存在する成分で、細胞構造の維持や抗原としての機能に関わっています。細菌は、ハンス・グラムによって考案されたグラム染色法によって、グラム陽性菌とグラム陰性菌に大別されます。グラム染色とは、紫色の色素を作用させてすべての細菌を染めた後、ヨウ素液による色素固定、アルコールによる脱色を行います。するとグラム陽性菌は厚いペプチドグリカン層を持つため、グラム染色に染まりますが、グラム陰性菌はペプチドグリカン層が薄く染まりにくいことからグラム陰性菌と呼ばれます。バクテリオファージが感染して食い尽くされてる大腸菌はグラム陰性細菌です。
どのようにしてバクテリオファージ(ファージ)が大腸菌に感染できるのでしょうか?バクテリオファージ(ファージ)が大腸菌に感染してその生活環を完成させるためには、感染の最初の過程である菌体表面への吸着を果たす必要があります。吸着はバクテリオファージ毎に特異的な宿主細菌に対してのみ生じるものであり、2つの段階を経て行われます。第1の過程は可逆的な結合であり、ファージのLTFが宿主細菌の持つ受容体に結合する。T4ファージはlong tail fiber (LTF) が大腸菌の細胞表面に存在するOmpCポリンタンパク質とリポ多糖 (LPS) に結合することで感染できるのです。OmpCポリンタンパク質は、グラム陰性菌である大腸菌の外膜に存在するタンパク質です。大腸菌の外膜には、さまざまなタンパク質から形成されるポリンと呼ばれるチャンネルが存在し、本来、薬剤が菌体内へ流入する役割を果たしています。
OmpCポリンタンパク質とはグラム陰性菌である大腸菌は、外膜と内膜で構成されており、これらに挟まれた空間をペリプラズムと呼びます。ペリプラズムとは、グラム陰性細菌の細胞膜と細胞外膜の間に存在する空間です。加水分解酵素や結合タンパク質などが存在し、物質代謝が発達したグラム陰性細菌もいます。外膜は、リポ多糖を主成分としており、抗菌薬が大腸菌体内へ流れ込む際の最初の防御壁となります。外膜は破壊されると内毒素(エンドトキシン)と呼ばれる有害物質を放出します。認識シグナルはLTF( long tail fiber)を通じて基盤に送られ、これによりshort tail fiber (STF) が不可逆的に大腸菌の細胞表面に結合する。認識シグナルとは、タンパク質のシグナル配列を認識して結合するシグナル認識粒子(SRP)のことでSRPは、タンパク質を合成しているリボソームと結合し、小胞体膜に存在するシグナル認識粒子受容体と結合することで、タンパク質を小胞体膜へと導きます。SRPは、タンパク質を合成しているリボソームと結合し、小胞体膜に存在するシグナル認識粒子受容体と結合することで、タンパク質を小胞体膜へと導きます。第2の過程である不可逆的過程においては、ファージの基盤がファージと細菌の結合を担うのです。
不可逆的な吸着を果たしたT4ファージは尾部の外側を覆う鞘を収縮させ、その内部に存在する管状構造(内筒)を細菌の細胞壁と細胞膜に向けて注入する。細胞壁にはペプチドグリカンの層が存在するが、先端のgp5がこれを分解する。注入された内筒は菌体の内膜を貫通せず、内膜と融合することでできた通路をファージのゲノムDNAが通過し、菌体内へと侵入する。ローリングサークル複製(Rolling Circle Replication)とは、環状のDNAを鋳型として、DNAの複製方法の1つで、DNAを複製する様式です。ローリングサークル複製では、DNAのリン酸と共有結合を作るタンパク質がレプリコンにコードされており、このタンパク質によって複製が開始されます。レプリコンとは単一の複製起点から複製されるDNA分子またはRNA分子、あるいはDNAまたはRNAの領域のことである。レプリコンワクチンとは自己増殖型mRNAワクチンのことです。
ローリングサークル複製は、生物が環境変化に適応したり、発生や分化を制御したりする際に利用されています。生物はヒストン修飾を利用した遺伝子増幅発現のスイッチのON/OFFにより、発現する遺伝子の数を爆発的に増やすことができます。
また、ローリングサークル増幅(Rolling circle amplification, RCA)という技術では、環状DNAを鋳型としてポリメラーゼ伸長反応によって長い一本鎖DNAを合成します。この技術は、細胞や組織中のDNAやRNAを検出する際に用いられます。ローリングサークル増幅の一本鎖DNAの合成の転写は通常の転写とは異なる様式でRNAを合成する反応です。ローリングサークル増幅反応(Rolling circle amplification, RCA)という手法もあります。これは、ローリングサークル増幅反応(Rolling circle amplification, RCA)、ローリングサークル複製とも呼ばれ英語でRolling Circle Replication略してRCRと書き、細胞や組織中のDNA・RNAの検出に用いられます。Replicationは複製という意味です。ウイロイドなどで確認されてRNA複製の仕方です。ウイロイド(Viroid)とは一本鎖環状RNA(250~400塩基)のみからなる最小の植物病原体でありウイロイドの RNAは,最も小さいウイルス RNAの 10分の1以下の大きさしかないが,その感染力はどの植物ウイルスよりもはるかに強いのです。RCR (Rolling Circle Replication)とはDNAの複製方法の1つで、ローリングサークル複製とも呼ばれます。ローリングサークル増幅(Rolling circle amplification, RCA)はローリングサークル増幅反応(Rolling circle amplification, RCA)と同じことで「環状DNAを鋳型としてポリメラーゼ伸長反応によって長い一本鎖DNAを合成」することです。
T4ファージの複製とパッケージングはT4ファージのゲノムは宿主のRolling Circle Replicationによって合成される。パッケージングとはバクテリオファージやウイルスの頭部に、DNAを取り込むことです。Rolling Circle Replication(ローリングサークル複製)にかかる時間はどれくらいでしょうか?ファージが大腸菌生菌の菌体内でDNA複製にかける時間は、ファージ感染大腸菌におけるDNA伸長速度として測定される。37°CでDNAの対数増加期における伸長速度は毎秒749bpである。DNA複製時における1塩基あたりの変異率は、1.7 × 10−8であり、T4ファージのDNA複製は非常に正確である。これは300コピーのT4ファージゲノムが、一つしかエラーを生じないことを意味する。また、T4ファージさえも独自のDNA修復機構を持っている。ファージの頭部は足場タンパク質の周囲に空の状態で組み立てられ、足場タンパク質はその後分解される。DNAは小さい孔を通過して前駆体頭部 (prohead) に格納されるが、DNAの格納に関わるのがDNAと最初相互作用するgp17(グリコプロテイン17)であり、この分子はDNAの折りたたみモーター及びヌクレアーゼとしても機能する。ヌクレアーゼ(nuclease)とは、核酸の糖とリン酸の間のホスホジエステル結合を加水分解してヌクレオチドにする酵素の総称です。核酸を分解する酵素として、DNAを分解するデオキシリボヌクレアーゼ(DNase)やRNAを分解するリボヌクレアーゼ(RNase)などがあります。ヌクレアーゼは、生体内ではDNA修復に関わる重要な機構です。
大腸菌の様な原核生物の免疫機構として、外来のウイルスDNAを分解して自己のDNAを守る機能があります。遺伝子工学では、DNAの塩基配列を認識して特異的に切断する制限酵素(Restriction enzyme)として用いられます。ヌクレアーゼには、核酸配列の末端から順に切断するエキソヌクレアーゼ(exonuclease)と、核酸配列の内部で核酸を切断するエンドヌクレアーゼ(nedonuclease)があります。
T4ファージの頭部にDNAを折りたたむ速度は1秒あたり2000塩基長であり、大きさを同等にした場合、その馬力は平均的な乗用車用エンジンに相当する。また大腸菌を破壊する溶菌サイクルにおいて、ファージのカプシドへ大腸菌のDNAの一部を取り込む形質導入が生じる。
T4ファージの放出は増殖過程の最終段階で、T4ファージは宿主となる菌体からウイルス粒子を放出する。ウイルス粒子の放出は細菌の細胞膜が破壊された後に起きる。ウイルスタンパク質がペプチドグリカンや細胞膜を破壊する溶菌を起こす。放出されたバクテリオファージは他の菌体に感染し、増殖サイクルを繰り返すのはヘルペスと似ています。ペプチドグリカンとはペプチドグリカンは、細菌の細胞壁を構成する高分子化合物で、糖とペプチドからできています。細胞壁の主要物質として、細胞の形態や強度を保持する役割を果たしています。
ペプチドグリカンとは、N-アセチルグルコサミンとN-アセチルムラミン酸が交互に連なった糖鎖に、ペプチドサブユニットが結合したポリマーです。ペプチドサブユニット同士が架橋されて網目状の構造をとっており、化学的に安定しています。ペプチドグリカンは、グラム陽性菌とグラム陰性菌の細胞壁に共通して存在しますが、構造が異なります。グラム陽性菌のペプチドグリカンはLys型、グラム陰性菌のペプチドグリカンはDAP型と呼ばれています。グラム陽性菌では細胞膜外に、グラム陰性菌では細胞外膜と内膜の間に存在しています。ペプチドグリカンは、酸やアルカリ、熱に強く不溶性ですが、リゾチームなどの酵素によって分解することができます。Herpesはリゾチームの酵素は持ってはいません。
部位特異的組み換えはヘルペスウイルスと同じように転移因子でも見られます。
転移因子とは何でしょうか?DNA型とRNA型に大きく分類されます。
DNA型:それ自身がゲノムDNAから切り出され、新しい部位に挿入されるcut-and-paste機構で転移します。
RNA型:RNA中間体を逆転写酵素によりDNAに変換し、ゲノムに挿入されるcopy-and-paste機構で転移します。RNA型はレトロトランスポゾンまたはレトロポゾンとも呼ばれ、ヒトゲノムに存在する転移因子の大半を占めています。
転移因子は、ゲノムのDNA配列を変化させることで突然変異の原因となり、多様性を増幅することで生物の進化を促進してきたと考えられています。また、遺伝学や分子生物学では、遺伝子を導入するためのベクターや、突然変異体を作製するための変異原として利用されています。
転移因子(トランスポゾン)とは、細胞内のゲノム上で位置を移動する(転移する)DNA配列を指し、動く遺伝子とも呼ばれます。ゲノム中の寄生配列と考えられていますが、進化の過程で遺伝子発現を制御するシス調節配列としての機能を獲得するなど、生物進化に貢献してきたと考えられています。
転移因子は、DNA型とRNA型に大きく分類されます。
DNA型:それ自身がゲノムDNAから切り出され、新しい部位に挿入されるcut-and-paste機構で転移します。
RNA型:RNA中間体を逆転写酵素によりDNAに変換し、ゲノムに挿入されるcopy-and-paste機構で転移します。RNA型はレトロトランスポゾンまたはレトロポゾンとも呼ばれ、ヒトゲノムに存在する転移因子の大半を占めています。
転移因子は、ゲノムのDNA配列を変化させることで突然変異の原因となり、多様性を増幅することで生物の進化を促進してきたと考えられています。また、遺伝学や分子生物学では、遺伝子を導入するためのベクターや、突然変異体を作製するための変異原として利用されています。
トランスポゾンとは、広義には細胞内でゲノム上の位置を移動する(転移する)DNA配列を指し、狭義にはその一種であるDNA(型)トランスポゾンを指す。前者は一般的に転移因子と呼ばれ、時にJumping genesとも呼ばれる。転移因子の中でも特にレトロポゾンはゲノム中で膨大なコピー数が存在し、例えばヒトゲノムの46%を占めるなど、ゲノム構造の多様性およびゲノムサイズに大きな影響を与えている。また、転移という特徴を利用し、遺伝学的ツールとして用いることができる。例えばP elementとよばれる転移因子を生体内で転移させることで変異体を多数作成し、特定の表現型を示す個体を選別してその原因となる転移因子の挿入サイトを特定することができる。ゲノムへの遺伝子導入への応用も広くおこなわれている。
転移因子の発見の歴史は転移因子の存在は、1950年にバーバラ・マクリントック(Barbara McClintock, 1902 – 1992年)によって提唱された。マクリントックは斑入りのトウモロコシの染色体を調べることで、斑の形成に関わる遺伝子座が染色体の異なる位置に移動することを発見し、この現象をトランスポジションと名付けた。しかし、遺伝子が染色体内を移動するといった概念は当時の常識をはるかに越えており、マクリントックの学説はほとんど受け入れられることは無かった。その後1960年代後半から70年代前半にかけて、バクテリアや酵母で「動く遺伝子」が発見されるようになり、それらはトランスポゾンと名付けられた。またマクリントックが発見したトウモロコシの斑の形成に関わる遺伝子座もDNAトランスポゾンの一種であることが明らかにされた。こうしてマクリントックが提唱した学説は認められるようになり、1983年には単独女性として初めてノーベル医学生理学賞を受賞した。
転移因子の分類は転移因子は、その転移機序に応じて2つのクラスに分類される。1つはカット&ペースト型の転移をするDNA(型)トランスポゾンであり、その転移機構をトランスポジションと呼ぶ。もう1つはレトロトランスポジション(またはレトロポジション)と呼ばれる転移機序を持つレトロトランスポゾンであり、いわゆるコピー&ペースト型の転移をおこなう。
またそれぞれの転移因子は、その転移様式によって自律型と非自律型に分類できる。自律型の転移因子は、その配列自身がコードする転移酵素(レトロトランスポゾンの場合は逆転写酵素)を利用して転移する。一方で非自律型の転移因子は転移のための酵素コードしておらず、自律型の転移因子から発現した酵素を利用して転移する。そのため、自律型のみならず非自律型の転移因子も、転移酵素や逆転写酵素の認識配列を持つのが一般的である。
DNAトランスポゾンやレトロトランスポゾンは、その転移機序と進化的起源によって細かく分類されている。特に最近は膨大なゲノム情報の蓄積によって、新規の転移因子ファミリーが次々と発見されており、中にはこれまでの分類に当てはまらない転移因子が発見されることもある。
DNA(型)トランスポゾンについてはクラスIIトランスポゾンとも呼ばれ、トランスポザーゼ(transposase)と呼ばれる転移酵素をコードしている。このトランスポザーゼがトランスポゾンの両末端にある逆向き配列(inverted repeat)を認識してゲノムから切り出し、切り出されたトランスポゾンDNAをゲノム上の別の位置に再び挿入させる。DNAトランスポゾンの種類によって、挿入のターゲットとなるゲノム配列の選好性は異なる。
DNAトランスポゾンは自律性と非自律性の2種類に分けられる。自律性トランスポゾンは上記のように内部にコードするトランスポザーゼを用いて転移する。一方で非自律性の因子は、内部にタンパク質をコードしておらず、自律性因子と類似した逆向き配列を持つ数百塩基対程度の短い因子である。一般にこうした短い非自律性トランスポゾンはminiature inverted-repeat transposable element(MITE)と呼ばれている。非自律性因子は自律性因子がコードするトランスポザーゼを利用して転移する。
レトロトランスポゾンについてはクラスIトランスポゾンとも呼ばれる。転移の際、まずレトロトランスポゾン配列が内部プロモーターによって転写され、自身がコードする逆転写酵素が翻訳される。逆転写酵素はレトロトランスポゾンRNAを鋳型としてcDNAを合成し、それをゲノム上の別の位置に挿入させる。したがってすべてのレトロポジションはレトロポゾン配列のコピー数の増加を伴い、ゲノムサイズに大きな影響を与えることが知られている。レトロトランスポゾンはlong terminal repeat(LTR)型レトロトランスポゾン、およびLINElong interspersed element(LINE)(非LTR型レトロトランスポゾンとも呼ばれる)に大別され、それぞれ次のように異なる転移機序を持つ。
LTR型レトロトランスポゾンは両末端に数百~数千塩基対の反復配列(LTR)を持ち、内部に逆転写酵素とインテグラーゼをコードしている。レトロウイルスはLTRに加えて内部にenv遺伝子を持つが、分類上LTR型レトロトランスポゾンに含まれることが多い。LTR型レトロトランスポゾンにもLTRのみを持つ非自律性因子が知られており、それらは長さに応じてLARDやTRIMと呼ばれることもある。
LINEは内部に逆転写酵素とエンドヌクレアーゼをコードする。LINEタンパク質はLINE RNAの3’末端配列を認識して結合し、逆転写およびゲノムへの挿入をおこなう。LINEには様々な種類が知られており、生物群ごとにそれぞれ異なる種類のLINEがゲノムの大きな割合を占める。例えば哺乳類ではL1と呼ばれるLINEが主要であり、例えばヒトゲノムの約17%はL1が占めている。一方、ニワトリゲノムではCR1、ゼブラフィッシュではL2がレトロトランスポゾンの中で最も大きな割合を占める。
さらにLINEに関連する非自律性因子として、short interspersed elements(SINE)が知られている。多くのSINEはtRNAに起源を持ち、RNA polymerase IIIによって転写されたSINE RNAが、LINEタンパク質を利用して転移する。そのためSINEの3’末端配列がLINEの3’末端配列と類似している例が数多く報告されている。SINEは進化的に異なる系統で独立に生成する場合が多く見られる。例えば霊長類のAlu、齧歯類のB1やB2など、哺乳類では目あるいは科レベルで独自のSINEを持つ。Aluはヒトゲノムの10%を占めており、そのコピー数は100万を超える。
転移因子のゲノムに対する影響については一般的に転移因子は、ゲノム中を移動するだけの利己的DNAあるいは寄生因子にすぎないと見なされる場合が多い。それでも、膨大なコピー数が存在し転移を繰り返していることから、その影響は無視できないものとなっている。例えば、転移因子が遺伝子内部に挿入されることで血友病や癌などの疾患をもたらす例が知られている。また複数の転移因子の間で非相同組換えが起こり、ゲノム構造を改変することもある。
転移因子の進化的視点から考えると、ゲノムサイズの増大、およびゲノム構造の多様化に大きな影響を与えており、ゲノムの重要な構成要素として認識されている。ゲノム中で転移因子の占める割合は、ヒトでは46%、トウモロコシでは80%以上であり、ゲノムサイズを決定する主要因となっている。一方で転移因子の水平伝播が起こっていることも知られているが、その詳細なメカニズムはほとんど明らかになっていない。
転移因子は上述のように有害な影響を及ぼす可能性があることから、一般の細胞内では転移が抑制されていることが多い。例外として、生殖細胞および脳神経細胞では転移が観察されており、特にニューロンのモザイク性を生み出している可能性が指摘されている。しかしなぜこれらの細胞内のみで転移可能なのかは明らかになっていない。体細胞における抑制機構としては、転移因子配列のメチル化やヘテロクロマチン化などのエピジェネティックな制御を受けていることが知られている。また、small RNAによる抑制も受けていることが近年明らかになってきている。
small RNAとは略してsRNAとは、遺伝子発現の制御因子として機能する代表的な分子で、原核生物から高等真核生物に至るまで、ほとんどの生物において存在しています。sRNAは、標的mRNAと塩基対を形成し、mRNAの翻訳や安定性を調節することで遺伝子発現を制御します。sRNAには、高等生物で見出されているmiRNA(micro-RNA)や細菌で見つかっているsRNAなどがあります。
転移因子の挿入は進化的に中立の変異であり、一般にその配列は進化の過程で塩基置換が蓄積し、やがて転移因子であることの検出が困難になると考えられる。しかし例外的に、その過程で転移因子由来の配列が生物の生存に有利な何らかの機能を獲得する場合が知られている(exaptationまたはco-optionとも呼ばれる)。それらの配列は進化的に保存されることが多く、特に有胎盤類では、保存領域の16%が転移因子に由来するとされているが有袋類ともいわれている。転移因子が獲得した機能としては、遺伝子のエキソン化や選択的スプライシングの生成といったタンパク質コード配列の改変に加え、エンハンサーやインシュレーターなどの調節配列となることが知られている。特に哺乳類の脳において複数のレトロポゾンがエンハンサー機能を有することが報告されており、哺乳類の脳の形成に関与したexaptationの好例として知られている。exaptation(エクザプテーション)とは、もともと備わっていなかった機能を得る過程、またはその過程によって発達した特徴を意味する英語です。
外適応(exaptation)の例としては、鳥類の羽毛が挙げられます。羽毛は現在では鳥が飛ぶために使われていますが、飛び始める何百万年も前には断熱材として使われていました。外適応とは元々備わっていなかった機能を得る過程とか外適応によって発達した特徴を言います。
転移因子の生物学的利用は、転移という特徴を利用し、転移因子を遺伝学的ツールとして用いることができる。具体的な利用方法の1つとして、転移因子を用いた順遺伝学的スクリーニングがある。これは転移因子を生体内で転移させることで変異体を多数作成し、特定の表現型を示す個体を選別してその原因となる転移因子の挿入サイトを特定する方法である。代表的なものとして、P element、Sleeping beauty、piggyBac、Tol2などが用いられている。この手法は、ショウジョウバエをはじめ、ゼブラフィッシュやマウスなどの脊椎動物でも幅広く用いられている。
また転移因子を利用したゲノムへの遺伝子導入、すなわち遺伝子組み換え生物の作成も広くおこなわれている。目的遺伝子の両端にDNAトランスポゾンの末端配列を付加したコンストラクト(構造物)を胚へ導入し、トランスポゼースを利用して転移させることで、効率良くゲノム中に導入することができる。この方法では、piggyBacやTol2がよく利用されている。トランスポザーゼ(トランスポゼース)とは、このトランスポザーゼという酵素は、トランスポゾンをゲノムから切り出す働きと、切り出されたトランスポゾンをゲノムに挿入する働きの両方を持っています。
ミトコンドリアとは何か?ミトコンドリアは、赤血球を除くすべての細胞に存在し、エネルギーの生産や細胞死、シグナル伝達など、生命活動を支える重要な器官です。ミトコンドリアの機能が低下すると、細胞が正しく働けなくなり、さまざまな臓器の機能に異常が起きるので癌はミトコンドリアのエネルギー(ATP)産生の低下で起こるという人もいます。その研究の一部を紹介します。
癌は複数の突然変異の蓄積により発生し進展すると考えられている.たとえば,あるひとつの上皮細胞は腫瘍原性の突然変異(がん遺伝子の活性化やがん抑制遺伝子の不活性化)を多段階的に蓄積し,さまざまな選択圧を克服しながら増殖能や浸潤能および転移能を獲得することによりがん化していくと考えられる.一方,最近になり,がんの発生および進展はこのような細胞の“自律的な”変異の蓄積のみによりひき起こされているのではなく,細胞どうしの相互作用を介した“非遺伝的な”変化も重要な役割を担っていることがわかってきた.たとえば,上皮に由来するがんの周辺には上皮細胞のほかにも間質細胞である線維芽細胞や免疫系細胞,炎症細胞などが存在するが,上皮のがん細胞はこれらの細胞と相互作用することによっても増殖能や浸潤能および転移能を獲得すると推察されている.また,ヒトのがん組織は必ずしも単一の細胞を起源とする遺伝的な背景をもつわけではなく,異なる遺伝的な背景をもつヘテロな細胞の集団であることもわかってきた.これらの事実は,異なる性質をもつ細胞どうしの相互作用が,がんの発生および進展に重要な役割をはたしている可能性を示唆している.しかし,そのような細胞間の相互作用がいかにしてがんの発生および進展に寄与しているのか,その分子機構はほとんどわかっていない。
何故そのような細胞間の相互作用がいかにしてがんの発生および進展に寄与しているのか,その分子機構はほとんどわかっていない根本的な唯一の理由として,癌は複数の突然変異の蓄積により発生し進展する生物の個体においてひき起こされたのではなくherpesウイルスがゲノムDNAの癌関連遺伝子(分裂増殖遺伝子)を変異させてしまったことが癌細胞(増殖過剰細胞)1個の誕生となります。しかも1個の癌細胞そのものは特別に恐ろしい細胞ではないのです。40兆個のたった1個の分裂増殖できる細胞が増殖し続けるためにはいくつかの条件を満たさなければならないのです。詳しくは後述します。
ミトコンドリアの機能障害とがんとの関係は古くから注目されてきた.これは,Warburgによる“がん細胞は正常な細胞とは異なり,酸素の濃度が十分な条件においてもミトコンドリアにおける酸化的リン酸化を使わず解糖系によりエネルギーを産生する”という観察(Warburg効果)に端を発する。Warburgは1950年代,この観察結果から,がん組織ではミトコンドリアに機能障害の起こっている可能性,さらには,ミトコンドリアの機能障害ががん化を駆動するという仮説(Warburg仮説)を提唱した。
このWarburg仮説の提唱から約60年の時をへて,実際に,がん組織においてミトコンドリアの機能障害が高頻度に認められることがわかってきた。とくに,ミトコンドリア呼吸鎖複合体の構成タンパク質をコードするミトコンドリアDNAには,膵臓がんにおいて高頻度に変異が見い出されているが,膵臓がんではRasシグナルの活性化も90%以上ときわめて高頻度に認められることが知られている。これは膵臓の細胞は自己増殖ができる特異的な細胞であるので膵臓の細胞に感染したヘルペスが膵臓の細胞の自己増殖するたびに大量のヘルペスが増殖することによって膵臓の細胞の数百個を超える増殖遺伝子(癌関連遺伝子)をものすごい速度で変異させてしまいます。しかもその変異を勝る勢いでヘルペスの数が増えてしまうのでますます膵臓の細胞の増殖遺伝子(癌関連遺伝子)が癌化(過剰増殖遺伝子化)させてしまうのですがその時に増えるヘルペスビリオンに正常な細胞のみならず癌細胞もすべての栄養素もエネルギーも奪われてしまい膵臓はherpesに占領されてしまうのです。増えすぎたヘルペスビリオンは膵臓だけで増殖することに満足できずに周辺の組織の細胞に荷の細簿も感染して新たな癌細胞を作り続ける転移病巣を作り続けてしまうのです。がんが恐ろしいのは癌細胞が増えることではなく正常細胞を癌細胞に変えてしまうヘルペスが無限に増えることです。
Warburg仮説と逆Warburg仮説のアポトーシスと内部共生説とは何か?
まず内部共生説とは何でしょうか?細胞内部共生説(symbiotic theory)は簡単に共生説とも言います。真核生物の細胞小器官は、異種生物がヒトの細胞内に取り込まれ細胞内で共生することにより生じたとする学説。細胞小器官であるミトコンドリアはα-プロテオバクテリア、葉緑体はシアノバクテリアが起源であり、核と細胞質の起源は古細菌であることが共通認識となっている。内部共生によって、新しい生物機能が創出されることが多く、動物や菌類、植物の祖先は、20億〜10億年前に起きた細胞内共生によって誕生したと考えられています。
真核生物の細胞小器官であるミトコンドリアや葉緑体も、異種の生物が細胞内に取り込まれて共生することで誕生したという仮説が「細胞内共生説」です。この説は、1970年にアメリカの生物学者リン・マーギュリスによって提唱されました。
細胞内共生説の根拠としては、次のようなことが挙げられます。ミトコンドリアは性質の異なる二重膜を持つ。ミトコンドリアは核ゲノムとは別にバクテリア由来の環状のゲノムを持つ。因みに癌の原因であるヘルペスウイルスビリオンが無数に分裂増殖すればするほど細胞は栄養素も核酸の原料もヘルペスに略奪され細胞小器官であるミトコンドリアも作れなくなりミトコンドリアの人体のエネルギー通貨であるATP産生能力もなくなっていくのです。その結果正常細胞も癌細胞も生き続けることができなくなり最終的には癌の原因であるヘルペスも死に絶えてしまうという皮肉な結果、すべてが死に絶えてしまうのです。
また、アリマキやシロアリ、アブラムシ、カメムシ、アズキゾウムシ、ウンカなど多くの種類の昆虫と微生物の細胞内共生が知られています。
Nobel prize winnerのOtto Warburgは,癌細胞が増えていけば好気性から嫌気性呼吸に典型的にスイッチすることを観察したのです。ミトコンドリア損傷が新生物(癌)形質転換を誘導すると仮定した。すぐ上で述べたようにヘルペスが増殖すればするほど細胞小器官のである一つに過ぎないミトコンドリアも栄養不足で指数関数的に減少してしまいます。その結果、いかなる癌においてもミトコンドリア機能低下は必ず起こります。
対照的に,病理学的老化は神経変性疾患におけるニューロン細胞において主に観察される。神経変性疾患の原因もヘルペスです。神経変性疾患についてはここを読んでください。ミトコンドリアにおける酸化呼吸によるATP産生はニューロンにおいて特に活性である。癌と神経変性疾患の間には逆共存症がある。これは,過剰なミトコンドリア活性が病理学的老化を誘導することにより,逆Warburg仮説が生まれましたが、ほんとうの逆Warburg仮説。の原因は神経線維細胞にはヘルペスは感染しないからです。何故ならば神経細胞は分れ列増殖することがほとんど無いからです。だから神経癌は存在しないのです。というのはヘルペスは分裂増殖をしない細胞には感染しないのです。
Warburg効果と逆Warburg仮説の両方が,酸化呼吸を介したアポトーシスの活性化または抑制により解明できることを示唆する。著者らの系統発生研究の重要な結果は,アポトーシスとアポトーシス様細胞死が,被食者プロトミトコンドリアと捕食者原始真核生物の間で行われた進化的アームレースにより進化したという発見であった。祖先のプロトミトコンドリア機械は毒性ミトコンドリア蛋白質を産生し放出する。現存するアポトーシス因子はこれらの毒素から進化した。著者らの実験は,ミトコンドリア機械が好気性条件への適応に直接関与することを示す。さらに,著者らの仮説は,異なるアポトーシス因子が呼吸に直接関与するという事実によって支持される。
ワールブルグ効果様代謝変化による抗腫瘍機能について説明します。
1. ワールブルグ効果
ドイツの生理学者であるオットー・ワールブルグ博士は,がん細胞が正常細胞とは異なるエネルギー代謝を行っていることを発見した1).酸素が十分に存在している条件下でも,ミトコンドリア活性を抑え,主に解糖系にグルコース代謝をシフトさせるというものである.この特性は「ワールブルグ効果」として認知され,ワールブルグ効果によるがん進展説は今もなお有力な作業仮説である.しかしながら,いまだにいくつかの点が解決されていない.たとえば,ワールブルグ効果は発がんの直接的なドライバー因子になりうるのか,もしくは悪性化したがん細胞の増殖や浸潤に必要な二次的な性状変化なのかについては,いまだ明確な答えが得られていない.一般的にワールブルグ効果はがん細胞の生存・増殖を助長すると考えられているが,がんの各ステージにおける腫瘍促進作用は正確にはわかっていない.「がんとは代謝疾患である」という概念も提唱されているなか,発がん初期におけるワールブルグ効果の意義を真に理解するためには,がん変異細胞の産生時に生じる代謝変化を詳細に検証する必要がある.本稿では,上皮層にがん細胞が出現したときのワールブルグ効果様代謝変化の意義について最新の知見を概説したい.
2. 細胞競合によりワールブルグ効果様代謝変化ががん変異細胞に生じ,上皮層より排除される。
正常上皮細胞層は抗腫瘍的な環境が整備しており,「細胞競合」と呼ばれる現象が重要な役割を担っていることが明らかとなってきている.細胞競合は,がん変異を有した細胞など個体にとって危険なものや環境に適応していない細胞を組織から排除することにより,生体内の恒常性維持に寄与する3).著者らの研究グループは,H-RasG12Vを発現した細胞(Ras変異細胞)と正常上皮細胞との細胞競合の結果,Ras変異細胞ではワールブルグ効果様の代謝変化が引き起こされ,管腔側に排除されることを見いだした4).ミトコンドリア内膜の電位差依存的に集積するtetramethylrhodamine methyl ester(TMRM)を用いてミトコンドリア活性を評価したところ,正常細胞に囲まれたRas変異細胞ではTMRMの集積が顕著に低下していた.一方,グルコースの細胞内への取り込みは促進し,乳酸脱水素酵素(lactate dehydrogenase A:LDHA)が細胞非自律的に発現亢進した結果,乳酸の産生量が増加した.Ras変異細胞を単独で培養したときにはこのような代謝変動は認められなかったことから,正常細胞と相互作用したRas変異細胞特異的にワールブルグ効果様の代謝変化が惹起されることがわかった.さらには,代謝関連酵素群の発現量を検討した結果,ピルビン酸脱水素酵素キナーゼ4(pyruvate dehydrogenase kinase 4:PDK4)が正常細胞と混合培養したRas変異細胞で増加することを突き止めた.PDK4はトリカルボン酸回路(tricarboxylic acid cycle:TCA cycle)のゲートキーパー分子であるピルビン酸脱水素酵素(pyruvate dehydrogenase:PDH)をリン酸化することにより,その活性を阻害する.すなわち,PDK4が過剰発現すると,ピルビン酸のTCA cycleへの流入が抑えられるためにミトコンドリアの機能が低下する.実際に,正常細胞に囲まれたRas変異細胞ではPDHが著しくリン酸化されており,PDK4を阻害すると,PDHのリン酸化は減弱しミトコンドリア機能が回復した.正常上皮細胞が変異細胞を管腔側に押し出す現象はepithelial defence against cancer(EDAC)と呼ばれ,アクチンフィラメントのクロスリンカーであるFilamin Aの集積がEDACに必須である5).一方,変異細胞内ではepithelial protein lost in neoplasm(EPLIN)がEDACのシグナルを下流分子に伝達することがわかっている6).Filamin AもしくはEPLINをノックダウンした,つまりEDACが不全な細胞ではPDK4の発現増加は認められず,ミトコンドリア活性も正常だったことから,EDACがワールブルグ効果様代謝変化を引き起こすことが明らかとなった.さらに重要なこととして,PDK4をノックアウトしたRas変異細胞を正常細胞と混合培養すると,管腔側への逸脱が顕著に抑制された.これらの結果より,EDACがPDK4を介した代謝変化を誘起させ,変異細胞を排除していることが示された.続いて,このような代謝制御によるがん変異細胞の排除が生体内で起きているかを検証するため,細胞競合マウスモデルを作出した.Cre依存的に活性化Ras変異を誘導でき,かつGFPでRas変異細胞を可視化できるLoxP-stop-LoxP-RasV12-ires-eGFPマウスを,腸管上皮特異的にCre-ERT2を発現するvillin-CreERT2マウスと掛け合わせた.タモキシフェンによるCre-ERT2の核内移行は確率論的に起きるため,タモキシフェンの投与量依存的に変異細胞の誘導率を調整できる.すなわち,低濃度のタモキシフェン添加下では少数のRas変異細胞が産生され,正常細胞とRas変異細胞との細胞競合を観察できる系を構築した.このマウスモデルの解析より,正常細胞に囲まれたRas変異細胞のほとんどが腸管の管腔へと排除されることがわかり,哺乳動物生体内において,細胞競合によりがん変異細胞が排除されることを世界で初めて実証した.また,マウス小腸陰窩のオルガノイドを用いてミトコンドリア活性を評価した結果,正常細胞に隣接するRas変異細胞ではTMRMシグナルが減弱することがわかった.さらに,iGT(intestine-specific gene transfer)法7)によりマウス小腸上皮細胞特異的にPDK4をノックダウンしたところ,Ras変異細胞の排除効率が有意に低下した.これらの結果より,マウス個体内においても,正常細胞に囲まれたRas変異細胞ではPDK4を介したワールブルグ効果様代謝変化が起こり,上皮層より排除されることがわかった.
3. EDACが誘導するワールブルグ効果と古典的ワールブルグ効果との違い。
これまで述べてきたように,EDACが誘導するワールブルグ効果様代謝変化はがん変異細胞を排除するのに対し,従来の古典的ワールブルグ効果はがん細胞の生存や浸潤を促進する.このように両者はがん進展においては相反する役割を担うが,グルコースの取り込み亢進や乳酸産生量の増加などの代謝特性は共通する.また,ワールブルグ博士は,ミトコンドリアの機能低下を代償するために解糖系が活性化すると考えており1),EDACによる代謝リプログラミングにおいてもミトコンドリアの脱分極が生じる.しかしながら,腫瘍細胞でもミトコンドリア機能は正常であるという報告が多数なされており,現在ではミトコンドリアの機能低下はワールブルグ効果を規定する指標の一つに必ずしも含まれていない8).分子機構の観点において,古典的なワールブルグ効果は,低酸素誘導因子1α(hypoxia inducible factor-1α:HIF-1α)が活性化され,下流のグルコース輸送体(glucose transporters:GLUTs),ヘキソキナーゼ1/2(hexokinase 1/2:HK1/2),LDHA, PDK1/3などを発現誘導することにより好気的解糖を促進する.一方,EDAC誘導型の代謝変化ではHIF-1αの活性化は認められず,PDK1/3ではなくPDK4が中心的な役割を果たしている4).PDK4の転写調節因子としてよく知られているのがペルオキシソーム増殖剤応答性受容体(peroxisome proliferator-activated receptor:PPAR)ファミリーである.大変興味深いことに,外部から細胞への圧縮刺激でPPARγの発現は変動し,EPLINもまた接着結合におけるアクチン重合を感知するメカノセンサーとして機能する.したがって,PPAR転写調節複合体がEDACに関与するかは現在のところ不明ではあるが,EDACによる物理的作用がワールブルグ効果様代謝変化を誘引する機序の一つであることが推察される.これが,低酸素や栄養枯渇などの腫瘍環境ストレスで誘引される古典的なワールブルグ効果の分子基盤との相違に起因すると考えられる(図1).

図1 EDAC誘導ワールブルグ効果と古典的ワールブルグ効果との違い
EDACが誘導するワールブルグ効果では,正常細胞からのEDACにより,変異細胞内でEPLIN→PDK4を介したワールブルグ効果様代謝変化が誘起され,上皮層より排除される.一方,古典的なワールブルグ効果は低酸素などの腫瘍環境ストレスによって引き起こされ,がん細胞の生存や浸潤を促進する.
4. EDACが誘導するワールブルグ効果の生物学的意義。
ワールブルグ効果によってグルコースを積極的に乳酸へと発酵することの生理的意義については,これまでにさまざまな議論がなされてきた.一つには,核酸やアミノ酸,脂肪酸などのバイオマスの合成量を増加させることにより,増殖が盛んながん細胞の代謝要求性を満たしているという解釈である.解糖系の中間代謝産物であるグルコース6-リン酸やフルクトース6-リン酸は核酸合成に用いられる一方,3-ホスホグリセリン酸やピルビン酸はアミノ酸の前駆体である.好気的解糖下では,ミトコンドリア内のアセチルCoAは細胞質へと排出され,脂肪酸合成にリサイクルされる.このような生合成経路の変動に加えて,ワールブルグ効果は細胞内の活性酸素種(reactive oxygen species:ROS)のバランスにも大きな影響を及ぼす.ワールブルグ効果によりペントースリン酸経路(pentose phosphate pathway:PPP)が活性化し,抗酸化剤である還元型ニコチンアミドアデニンジヌクレオチドリン酸(nicotinamide adenine dinucleotide phosphate, reduced:NADPH)の産生を促す.また,ミトコンドリアは主要なROS発生器官であるため,がん細胞は自発的にミトコンドリアを不活性化させ,細胞増殖の副産物であるROSのレベルを低下させることも提唱されている.よって,細胞競合下で観察されるミトコンドリアの機能低下は,EDACによる酸化ストレスへの耐性獲得に寄与しているのかもしれない.しかしながら最近の研究成果によると,TCA cycleの酵素であるイソクエン酸デヒドロゲナーゼ2(isocitrate dehydrogenase 2:IDH2)がNADPHを産生することで,ミトコンドリアはROS産生に拮抗的に働くことが示唆されている.また,ROSはDNA障害を引き起こすばかりでなく,シグナル伝達因子としても機能する.具体的には,ROSがphosphatase and tensin homolog(PTEN)やチロシンホスファターゼなどの脱リン酸化酵素を不活性化することが知られており,EDACシグナルを仲介するメディエーターとしてROSが機能する可能性も考えられる.このような点からも,細胞競合時のがん変異細胞がどのようなレドックス状態になっているかは大変興味深く,今後詳細に検討する必要がある.ミトコンドリアの電子伝達系では36分子のATPが産生されるのに対し,解糖では2分子のATPしか産生されない.このように解糖は一見効率が悪いが,ATPの合成速度に着目すると,呼吸鎖に比べて解糖の方が100倍ほど早い.つまり,細胞外にグルコースが豊富にある環境では,積極的に解糖系で代謝することによりATPが過剰に産生される.実際に,FRETプローブを用いた結果より,正常細胞と共培養したRas変異細胞のATPレベルは単独培養時に比べて有意に増加していた4).この結果から,ミトコンドリア代謝から解糖へのシフトは,局時的にATP産生量を増加させ,変異細胞が上皮層より逸脱するために必要な細胞内活動を支持することが示唆された(図2).

図2 EDACが誘導するワールブルグ効果様代謝変化の生物学的意義
正常細胞に隣接する変異細胞は,ミトコンドリアの機能低下に伴って解糖が亢進する.この代謝シフトによって細胞内ATP量は増加するが,ROSのレベルがどう変化するかは現在のところ明らかではない.
5. まとめ
がんの進展を促進するという従来のワールブルグ効果とは対照的に,がん細胞が産生されたときに生じるワールブルグ効果様の代謝変化は,むしろ発がんに抑制的に機能することが示された.分子レベルでの相違点はあるものの,代謝特性は非常に類似していることから,ワールブルグ効果の多様な役割が明らかとなってきた.今後,がんの各ステージで特異的に機能する代謝関連因子を同定することによって,代謝をターゲットとしたがん治療法のブレークスルーになることを期待したい.
選択的スプライシングと,がんのワールブルグ効果。
解糖系酵素ピルビン酸キナーゼM(PKM)には構成的活性型のPKM1, 条件的活性化型のPKM2という二つのスプライシングアイソフォームが存在し,大半のがんがPKM2を選択的に発現する.かつて,PKM2はワールブルグ効果の形成を通じて腫瘍細胞に代謝上の有利をもたらすとされたが,PKM2欠損マウスの表現型は“がん促進”だった.筆者らは,Pkmの選択的スプライシング制御を固定化した新たな遺伝子改変マウス群を作製し,がんや代謝制御におけるPKM1・PKM2の役割を再訪した.マウス発がん試験や細胞移植実験などによって,真に腫瘍促進的なのは,PKM2ではなく,むしろPKM1の方であることがわかった.実際にPKM1を高発現し,その生存・増殖をPKM1に依存する高悪性がんの存在も明らかになってきた.
1. はじめに
活発に増殖を繰り返す腫瘍細胞は,自身を形作る細胞成分等を合成し続ける必要がある.また,それを可能にするため,高エネルギー状態を維持することも重要となる.そのような需要に合わせ,がんは特徴的な代謝形質を持つことが知られる1).がんの代謝特性として最もよく知られるものの一つが,ワールブルグ効果だろう.知名度が非常に高いせいか,逆に,人によって定義がまちまちだったりする問題があって悩ましい.本稿では,発見者であるO. Warburg自身の観察結果に即し,「グルコース取り込みの著しい亢進,乳酸産生・分泌の亢進」をワールブルグ効果とする.
わざわざ断りを入れたのには理由がある.たとえばワールブルグ効果を「(好気的)解糖系の亢進」と表現する人もいる.その言い回しは間違いではないが,ややあいまいで誤解を招きかねないので筆者は避けている.理由はいくつかあるが,最大のものは,がんと好気呼吸にまつわる問題との兼ね合いである.事実として,がんでは解糖系・ミトコンドリア好気呼吸の両方が亢進している.かつては,「がんでは好気呼吸が抑制されている」と誤解されていたと聞く.ワールブルグ自身も「想像」として,そのように語ったことがあったようだが3),実は,もともとのワールブルグ効果の定義に「好気呼吸の抑制」は入っていない.
むしろ,近年,がんにおけるミトコンドリア好気呼吸の亢進およびその意義が,疑いなく示された4–8).これは,当該研究分野における,この10~15年で最大の成果の一つといっていいほどのものだ.しかるに見わたすと,「がんではTCA/酸化的リン酸化が“抑制”されている」というストーリーを信奉する人が,少数ながら,いまだいるように見受けられる.早く誤解が解けることを願っている.またそのような人は「乳酸産生=解糖系活性」と考えてしまう傾向があるように感じられるが,事はそう簡単ではないとお伝えしたい.解糖系を経由するグルコースの代謝先は,乳酸だけではない.また,反対に,乳酸のソースはグルコースだけではないということもある(アミノ酸等からも乳酸が作られている).
ともかく2010年代の前半ごろにワールブルグ効果の一大ブームが起きたとき,今振り返ると,がん代謝の研究分野は非常に偏ったコンセプトに満ちていた.たとえば,“細胞増殖にはワールブルグ効果が必須”,“ミトコンドリア好気呼吸は細胞増殖にネガティブ”といった類のものである.それらミスリードの起点となったのが,解糖系酵素ピルビン酸キナーゼM(PKM)に関する誤解だった,と筆者は考えている.本稿では,筆者らの研究を中心に,関連分野の歴史も含めて概説する.
2. PKMのスプライシングアイソフォーム
PKM(pyruvate kinase M)は解糖系の後半部にて,ホスホエノールピルビン酸をピルビン酸へと変換する.この反応は,生理的条件下では不可逆反応といってよく,ゆえに解糖系にいくつか存在する不可逆ステップの一つとなっている.PKMにはPKM1, PKM2という二つのスプライシングアイソフォームが存在する(図1A).両者はエクソン9,エクソン10という特異的で相互排他的なエクソンのどちらがmRNA上に残されるかが異なっている.そのような選択的スプライシングの結果,活性制御が異なるタンパク質が生じるという点がとてもユニークである.PKM1は自発的に活性型の四量体を形成することができる.いわば構成的活性化型である.これに対し,PKM2が四量体化するためにはアロステリック因子が必要で,条件的活性化型といえる.アロステリック因子とはタンパク質の活性部位以外の部位に結合することで、そのタンパク質の活性や機能を調整する化合物のことアロステリック因子がない場合,PKM2のKm値(PEPに対する)は10倍以上高く,Vmaxも半分程度で,その活性は非常に低い9).PKM2のアロステリック活性化因子は複数同定されているが10),最も代表的なのはFBPで,これは解糖系上流の中間産物である(図1B).結果的に,PKM2は,解糖系上流の代謝物レベルが一定以上になったときだけ活性化する状況になっている.したがい,構成的活性化型のPKM1と比べた場合,PKM2にはグルコースからピルビン酸へのフラックスを低く保つ作用がある.

図1 PKMのスプライシングアイソフォーム
(A)選択的スプライシングの模式図.ボックス内の数字はエクソン番号.(B) PKM2特異的なアロステリック制御機構.FBP:fructose 1,6-bisphosphate, PEP:phosphoenolpyruvate, Pyr:pyruvate, Lac:lactate.
PKMの選択的スプライシング制御は,基本的に相互排他的になっている.それに関し,かつて,「PKM1は成人型,PKM2は胎児型」と呼ばれた時代もあったが,それらは実態を反映したものではないことが現在はわかっている.また,細胞周期も含め,細胞の増殖状態とことさらに関連しているわけではない.「正常細胞はPKM1を,腫瘍細胞はPKM2を発現」などの捉え方は,暴論の極みだ.
むしろ,筆者らのものを含むいくつかの報告から考えると,PKMのスプライシング制御は,おそらく,細胞分化との関連性が最も高い8, 11, 12).具体的には,成人であっても,PKM1発現は,ごく一部の細胞タイプに限定されていて,筋分化や神経分化との相関をうかがわせる(図2).それは胎児期であっても同様である(未発表).一方,それ以外の広範なタイプの細胞は,PKM2発現型であることが多い(肝細胞のように,どちらも発現しないタイプも存在する).大半の上皮細胞,神経幹細胞,グリア,線維芽細胞等がPKM2型に該当する.ヒトで発生するがんの多くが,それらもともとPKM2型の細胞を起源としていることに注意してほしい.よくいわれるように,ほぼすべてのがんがPKM2を発現しているのは事実だ.しかしそれは,一般論で言って,別に細胞のがん化に伴って獲得される性質ではない.起源細胞の段階から,PKM1を発現していないことに注意しなければならない.

図2 各種正常細胞におけるPKMの発現アイソフォーム
3. PKM2/ワールブルグ効果と,がん
ほぼすべてのがんがPKM2を発現することは古くから知られていたが,その現象の注目度を一気に高めたのが,腫瘍細胞のPKM2発現とワールブルグ効果を関連づけたVander Heidenらのグループの研究である.彼らは,肺がん細胞株H1299に外来性PKM1またはPKM2を発現させた上で内在性PKM(主にPKM2)をノックダウンし,PKM1またはPKM2のどちらか一方のみを発現する細胞を作製した(図3A)。表現型解析の結果,低酸素条件での増殖やマウス移植モデルでの腫瘍形成に,PKM2発現型の方が有利という結果が報告されている.代謝面では,PKM1発現細胞よりも,PKM2発現細胞の方が乳酸産生が高かった.一連の結果から,「ワールブルグ効果様の代謝にはPKM2発現が重要(PKM1はその作用に拮抗)で,それによって低酸素下での増殖や腫瘍形成が促進される」というモデルが提唱された.ワールブルグ効果の意義を実験的に初めて示した(ようにみえる)研究で,きわめて大きなインパクトがあった.冒頭付近で述べた,ワールブルグ効果フィーバーは,おそらくこの報告を起点としている.

図3 「PKM2/ワールブルグ効果はがんに有利」仮説の始まりと,終わりの始まりを示す。
(A)外来性Pkm過剰発現と内在性PKMノックダウンを組み合わせた,PKM1/PKM2片方のみを発現する細胞株の作製.仮説の始まりを示す。
.(B) Pkm2ノックアウトマウスにおける遺伝子改変.仮説の終わりの始まりを示す。ノックダウンとは遺伝子の活動を部分的に減少させる遺伝子改変技術でノックアウトマウスとは、遺伝子操作によって特定の遺伝子を欠損(無効化)させたマウスです。遺伝子の塩基配列は決定しているものの、その遺伝子産物の機能が不明な場合に、その機能を推定するための重要なモデル動物として利用されています。
ところが数年後,同グループにて作製されたPkm2欠損マウス(Cre-loxP系を用いたコンディショナルKO,図3B)が,まったく想定外の表現型を示した.“がんを促進するはずのPkm2をノックアウト”したところ,複数実験系にて,“発がんやがん進展が促進”されてしまったのである14–16).この段階で前述の「PKM2/ワールブルグ効果は,がんに有利」モデルは,少なくとも,一般性を失ったようにみえた.詳細は省くが,上記表現型の説明として,「PKM活性が高いと(PKM1の発現はこれに該当する)細胞増殖が止まってしまう」という代替説が披見された14, 17).しかし,自らの先行研究における実験結果との矛盾,また,かなりアクロバティックな理屈によるモデル構築に疑問の声(表には出にくいが)もあった.
このころから,「ワールブルグ効果はがんに有利」仮説を否定するような重要反証が,少数派ながら,いくつかあがるようになっていた4, 5).一方で,上記仮説を無条件の前提として開始された多くの研究・論文(あまり質が高いとはいえない)の量産は,むしろ全盛期に差し掛かろうとしていた.振り返ると,筆者が直接に見聞きした範囲で,がん代謝の分野が近年で最も混沌としたのがこのころだったように思える.ちなみに上記マウス系統をもとに全身性の恒常的Pkm2-KOマウス(ストレートKOマウス)も作製され,正常細胞の増殖や発生にもPKM2が必須でないことが確認されている.
4. ノックインモデルによる,PKM1がん促進的機能の発見。ノックインモデルとは遺伝子編集技術によって特定の遺伝子領域に任意の遺伝子配列を導入した細胞やマウスなどのモデル生物を指します。通常,ノックアウトのような遺伝子改変マウスでの実験結果は,手法がシンプルなだけに,関連分野の混乱を収める決定打になることが多い.しかし,PKM2の場合にはそうならなかった.その理由は,PKMにスプライシングアイソフォームが存在するからかもしれない.Pkm2ノックアウトマウスでは,本当にシンプルに,PKM2特異的エクソンのみがゲノムから削られた.しかし,そのような遺伝子改変の結果起きることは,むしろ複雑である.まず,crypticスプライシングが起きたようだが,生じるmRNAがコードするのは酵素活性を持たないタンパク質なのであまり問題にはならない.一方で,一部にPKM1型エクソンをピックしたmRNAが作られ,PKM2消失の代わりにPKM1が作られるようになった.一番の問題は,このPKM1へのスイッチが,中途半端かつコントロール不能に起きたことにある.ここに,さらに,Cre-loxPによる組換え効率は100%ではないことが加わって状況がいっそう複雑化した14).
それら問題を解決し,PKMスプライシングアイソフォームそれぞれの機能を評価しようと,筆者らは新たな遺伝子改変マウスを開発した.選択的スプライシングの切り替えを不能化することを主コンセプトに,Pkmノックインマウスを作製した.これらマウスでは,内在性Pkm遺伝子のエクソン8途中に,PKM1またはPKM2のcDNA後半部分(と,ポリA付加シグナル)をシームレスに挿入してある(図4).その結果,PkmM1アリルからはPKM1のみが,PkmM2アリルからはPKM2のみが作られる.その結果,ホモ変異マウスでは,発現量はほぼ等しいが,アイソフォームはPKM1まはたPKM2のどちらかのみ,という状況を作ることができた.転写自体は内在性プロモーターによっているため,発現量や発現細胞の特異性も,野生型のそれをほぼ完全に再現している9).これまでのところ,PKM1またはPKM2どちらのホモノックインマウスも,発生,生育,繁殖等にきわだった異常は出ていない.したがい,どちらかのアイソフォームが発現していれば,マウスはほぼ正常を保てると考えられる.一方で,PKM1およびPKM2の両方を欠損させたマウスは胚性致死である(未発表).

図4 Pkmノックインマウスにおける遺伝子改変の模式図
エクソン8内の↓位置に,PKM1またはPKM2 cDNA後半部分を,ポリA付加シグナルとともにシームレスに挿入(ノックイン)してある.
我々は,Pkmノックインマウスを用い,活性変異型Kras誘導性の肺発がん実験(肺腺がんを誘導),発がんイニシエーターであるDMBAを用いた化学発がん実験(皮膚がん,肝臓がん,肺がん,リンパ腫,等を誘導)を行った.それら実験の結果から,PKM1に,PKM2よりも強い発がんプロモーター活性があることがわかった9).一般的には,真にがん促進的なのは,PKM2よりも,むしろPKM1であることが強く示唆された.また,それらマウス由来の細胞を用いた形質転換実験・腫瘍形成実験等の結果から,PKM1によるがん促進作用は,細胞自律的な機序によることがわかった.前述Pkm2ノックアウトマウスがみせた“がん促進”の表現型は,代償的に発現したPKM1の作用によって説明できると考察された.
5. “PKM1 vs PKM2”によるグルコースおよびグルタミン代謝の制御
PKM1による“がん促進”の機序を明らかにするため,ノックインマウス由来細胞を用いた代謝解析を行った.PKM1は構成的活性化型である一方,PKM2の活性はアロステリック制御の下にある.ゆえに大雑把には,PKM1は超高活性型,PKM2は低活性型といって構わないだろうことはすでに述べた.
まず,安定同位体標識グルコースを用いたトレーサー解析の結果から,PKM1が,PKM2と比べ,グルコース代謝を亢進させることがわかった.グルコースに由来する乳酸産生,グルコースからTCA回路への代謝フラックス,そのいずれもがPKM1によって亢進していた.従前の仮説として,PKM1の高活性によってグルコースからTCA回路へのフラックスが高まると,同じくグルコースを栄養源とするペントースリン酸経路(核酸合成やNADPH産生に寄与),プリン合成,セリン合成に支障が出るのでは?と想像されてきた14).しかし,実際には,少なくともそれら代謝経路に問題は出ないことが,トレーサー解析,メタボローム解析の結果から明らかになった.すなわち,PKM1はグルコース代謝を同化,異化の両面で亢進させ,それが腫瘍に種々の代謝メリットをもたらすことが強く示唆された.
一方で非常に興味深かったのが,“総”乳酸産生の問題である.すでに述べたとおり,PKM1は,PKM2と比べ,グルコースから乳酸への変換を亢進させる.また,短期的には,総乳酸産生も,PKM1細胞の方が高い.しかし,やや長期の観察期間でみた場合,PKM2細胞の方が,PKM1細胞よりも多くの乳酸を産生していた.この現象は,Cristofkらが観察した結果とよく似ている13).一連の結果は,PKM2発現下における乳酸産生に,グルコース以外のものが含まれていることを示唆していた.
種々の状況証拠や先行研究から,グルタミン(増殖細胞にとって,グルコースと並ぶ重要な炭素源)代謝の関与が考えられた.そこで,安定同位体グルタミンを用いたトレーサー解析を行ったところ,PKM2細胞では(PKM1細胞と比べ),グルコースからTCA回路へのフラックスが低下している代わりに,グルタミンからTCA回路へのフラックスが高いことがわかった9).TCA回路に入ったグルタミンは,おそらくリンゴ酸酵素ME1/2の働き等によって,最終的に乳酸へと変換されていた(図5).これら結果から,PKM2発現細胞でみられる高い乳酸産生は,グルタミン由来乳酸の増加によるものだろうと考えられた.個人的教訓として,中心炭素代謝全体が非常に柔軟なネットワーク構造として存在していることを再認識させられた.

図5 グルタミンを栄養源とするTCAサイクルの回転と乳酸産生
TCA:TCAサイクル,PPP:pentose-phosphate pathway.
6. 小細胞肺がん,PKM1依存の発見
ここまでの我々の結果から,超高活性型であるPKM1は,もし発現すればPKM2以上の代謝メリットを腫瘍にもたらすと予測された.しかし,PKM1を発現するがんはヒト,動物含めてまったく知られていなかった.筆者らは,その理由を,がん起源細胞の問題に結びつけて考察した.
そして,肺・気管支に存在する神経内分泌細胞に注目した.この細胞は,小細胞肺がんという肺がんサブタイプの起源細胞と想定されている.小細胞肺がんは,肺がんの中でも予後が悪く,他がんと比べて治療標的の開拓が遅れている難治がんである.ノックインマウスにおけるPKM1, PKM2発現の組織特異性を注意深く調べる過程で,筆者らは,肺(正確には気管支・細気管支)の神経内分泌細胞が,PKM1陽性かつPKM2陰性(ほぼ検出できないレベル)であることに気づいていた9).この細胞を起源とする小細胞肺がんは,PKM1を発現している可能性が高いと考えた.
予想は正しく,細胞株パネル・臨床検体を使った解析によって,他がん種とは明らかにレベルの違う高PKM1発現を小細胞肺がんにみとめた.ただし,PKM1が優勢というところまではいっておらず,質量分析によるタンパク質レベル絶対定量の結果からは,PKM2が全PKMの60~70%程度を占めていることがわかった.正常神経内分泌細胞と比べてPKM2レベルが高いのは,おそらく,小細胞肺がんではMYCファミリー転写因子の活性が非常に強いためと思われる(後述).しかし,この程度までPKM1の割合が高まると,マニアックで面白いことが起きる.あまり知られていないが,PKM1は,アロステリック因子なしにPKM2とのヘテロ四量体を形成する(正確なサブユニット比は不明)(図6)9).すなわち,PKM1には,PKM2を強制的に活性化する能力がある.実際,Blue-Native PAGE等の生化学的解析によって,小細胞肺がん細胞株で発現するPKM2が(おそらくPKM1の作用によって)アロステリック因子なしに四量体形成していることが確認された9).まとめると,小細胞肺がんはPKM1とPKM2とが混在する珍しい状態にあるがんだが,そのPKM活性制御は,ほぼPKM1型になっている。アロステリック因子とはアロステリック因子とは、タンパク質の活性部位以外の部位に結合することで、そのタンパク質の活性や機能を調整する化合物のことを指します。アロステリック因子によってタンパク質の立体構造が変化することで、その活性や機能が調整されます。

図6 活性型PKM1–PKM2ヘテロ四量体の形成
通常のPKM2四量体形成にはアロステリック因子が必要だが(上),PKM1–PKM2ヘテロ四量体形成には不要(下).M1:PKM1, M2:PKM2.
小細胞肺がんにおけるPKM1発現の意義を調べるため,ヒト小細胞肺がん細胞株に外来性PKM1またはPKM2を発現させた上で,内在性PKM(PKM1, PKM2の両方)をRNAiによってノックダウンして表現型を観察した.その結果,小細胞肺がん細胞の生存や増殖にはPKM1が必須であり,PKM2によっては代替できないことがわかった。PKM1がもたらす高グルコース代謝が,このがんにとって不可欠な要素となっていることが示唆された.
7. 神経内分泌がんとPKM1発現
PKM1を発現する小細胞肺がんが示す特徴の一つとして,神経内分泌分化の形質がある.正常な神経内分泌細胞がPKM1型だったことと合わせ,PKM1発現と神経内分泌分化がリンクする可能性が高いと考えた.この仮説の妥当性を計るため,筆者らは次に,前立腺がんに着目した.前立腺小細胞がんは,神経内分泌分化の点を含め,小細胞肺がんとの類似性が非常に高いがんとされる20).de novoで発生する前立腺小細胞がんはそれほど多くないが,近年,去勢抵抗性前立腺がん(男性ホルモンであるアンドロゲンをターゲットとする内分泌治療の過程で耐性化した再発がん)の多くが神経内分泌系列にlineage転換していることが明らかになっている20, 21).前立腺がん細胞株パネルで検討すると,神経内分泌分化形質を示す株は,腺がんタイプのものと比べてPKM1/PKM2 mRNA比が高いことがわかった(図7A).

図7 神経内分泌がんとPKM1
(A)前立腺がん細胞株パネルでの解析.AdPC:前立腺腺がん,CRPC:去勢抵抗性前立腺がん,SCPC:前立腺小細胞がん,SCLC:小細胞肺がん,AR:アンドロゲン受容体,NE:神経内分泌分化.(B)神経内分泌分化を誘導できるヒトオルガノイド.DOX:ドキシサイクリン.
PKM1と神経内分泌分化の関係をより直接的に調べるため,Tet-Onシステムによって神経内分泌分化を誘導できるヒトオルガノイド株TR-6TF22)を用いて検討を行った(図7B).このオルガノイドはヒト正常大腸由来で,ゲノム編集によってTP53およびRB1遺伝子(それぞれp53, pRBをコード)を不活化した(両遺伝子の同時機能喪失が神経内分泌分化がんの特徴のため)のち,神経内分泌分化に関連すると予想される六つの転写因子(ASCL1, SOX2, NKX2.5, TP73, NEURODO1, POU3F2)を遺伝子導入(Tet-ONによる誘導発現)してある.ドキシサイクリン添加の前後で遺伝子発現解析を行ったところ,分化後のPKM1/PKM2 mRNA比上昇がみとめられた(未発表).
一連の結果から,肺がんに限らず,神経内分泌分化がん一般が広くPKM1を発現している可能性が高まっており,現在はさらに,種々の消化器臓器に由来するがんで検討を行っている.
8. PKMの選択的スプライシング制御機構
これまでに,PTBファミリーに属する複数のスプライシング因子が,PKMの選択的スプライシング制御因子として同定されている.具体的には,PTBP1(PTB),HNRNPA1, HNRNPA2等である.これらスプライシング因子の発現は,MYCファミリー転写因子の制御下にあって,PKM1型スプライシングの抑制に寄与している.MYCファミリーの下流ということで,細胞の増殖状態とPKMのスプライシング制御に関連性があるようにも思える.しかし,筆者らの成績では,細胞を静止期に導入(血清飢餓または接触阻害にて)してもPKM1/PKM2比はほとんど影響を受けないということもあり(未発表),それほど単純な話ではないのだろうと思われる.一方で,細胞老化を誘導した場合には,明らかにPKM1/PKM2比が上昇する(図8).また,別のスプライシング因子であるSRSF3が,PKM2型スプライシングを促進する因子として同定されている.PKM1型スプライシングを積極的に促進するスプライシング因子として報告されたものはいまだないが,存在する可能性は高いと考えている.その場合,おそらくその因子は,神経・筋・神経内分泌分化等に関連するものだろうと予想している.

図8 細胞老化に伴うPKM2からPKM1へのスイッチ
(A)初代培養MEFの「継代による老化・不死化」過程の解析.(B)がん遺伝子誘導性細胞老化(OIS:oncogene-induced cellular senescence)での解析.正常細胞(この場合は初代培養MEF)に,p53不活化等による不死化を経ないまま,活性化型Rasを発現させると細胞老化が誘導される.(C)治療誘導性細胞老化での解析.Kras形質転換MEFをDNA障害性抗がん剤であるシスプラチンで処理すると,治療誘導性の細胞老化が誘導される.
9. おわりに
本稿ではPKMスプライシングアイソフォームについて,現時点までの研究の変遷を含めて概説した.本酵素は特に,ワールブルグ効果との間の因縁が深いため,その点から起稿した.がん代謝の分野を一時席巻した「ワールブルグ効果=がんに有利」仮説について,少なくともPKM2研究の結果にもとづく部分については,ほぼ振り出しに戻ったような状況と捉えている.それ以外の研究の成果も含め,現在,ワールブルグ効果の意義を問われれば「よくわかっていない」と答えるのが正しい,というのが世界的主流のように思える.その文脈では,最近,乳酸について,がんにとって排出物というよりもむしろ栄養源である可能性や,腫瘍免役との相互作用も指摘されており,興味深い.
ちなみに筆者らは,腫瘍にとって多くの代謝メリットを得られるのは,一般的にはPKM1の方だと考えているが,確かに,まれにPKM2の方を好む株が存在することも経験している(未発表).おそらくH1299細胞もそのような株の一つだったのではないかと想像している.PKM1とPKM2,どちらを好むか? その規定因子のようなものが明らかになれば,一連の齟齬は完全に解消されるだろう.
PKM1/PKM2の問題は,単一遺伝子の選択的スプライシングが,エネルギー代謝のフレームワークや細胞(小細胞肺がん等)によってはその生死を決定しうる珍しい例かもしれない.同時に,発がん実験等を行わない限りノックインマウスに目立った表現型異常が出ていないことから,生理的条件下における代謝ネットワークの高い柔軟性をうかがうことができる.筆者らは現在,主に,PKM1がもたらす代謝メリットをより詳細に調べ,神経内分泌がんの新規治療標的として開拓する試みを行っている.一方,もしPKM1型スプライシングを促進するRNA結合タンパク等がわかれば,同様に新規治療標的になる可能性がある.以前はスプライシング因子に対する阻害剤開発というと相当なハードルの高さを感じたが,PROTAC(proteolysis targeting chimera)等の新技術を活用して実現する途を探ろうとしている.
がんの健診でPETと言う言葉を聞いたことがあるかもしれません。これは、「がん細胞がブドウ糖/グルコースを多量に取り込む」という特性を利用して、目に見えないがん細胞の塊(腫瘍)を見つける検査法です。
がん細胞は正常細胞の何倍もの量のグルコースを取り込むので、グルコースと同じ挙動を取る18F-FDG(放射性物質)を注射するとがんの病巣に集まります。集まったところからは放射線が放出されるので、それを捕らえて画像化してがんの病巣を見つけ出すことができます。
この「がん細胞がブドウ糖(グルコース)を多量に取り込む」という性質は、100年ほど前にノーベル賞受賞者のOtto Warburgが観察した現象で、Warburg(ワールブルグ)効果と呼ばれます。がん細胞は有酸素下でも効率の良いミトコンドリアの酸化的リン酸化よりも、効率の極めて悪い解糖系で生物がエネルギー源として使うATPを産生するのです。グルコースは解糖系で代謝された後にミトコンドリアに入ることなくピルビン酸キナーゼ(PKM)によってピルビン酸に変換され、がん細胞は大量のグルコースを消費することになります(この辺は高校の生物の教科書に詳しく書いてあります)。これらのATP産生の切り替えに係るPKMですが、1つの遺伝子から2つの性質が異なるPKM1とPKM2がmRNAの選択的スプライシングという現象によって生み出されることが判明していました。
1つの遺伝子から2つの性質が異なるPKM1とPKM2がmRNAの選択的スプライシングという現象によって生み出される。PKM2がワールブルグ効果の原因とされる。
PKM1は、グルコース(Glucose:赤)由来炭素のTCA回路への流入と乳酸(Lac)への変換のいずれをも促進させる(グルコース消費を亢進)。代謝ネットワークの変化が、未解明の制御機構を通じて総乳酸産生を低下させる。PKM2は、グルタミン(Gln:緑)消費を亢進させ、乳酸産生が増加する(グルタミン消費を亢進)。
ミトコンドリアの機能低下に関係する病気の総称を「ミトコンドリア病」といい、遺伝子の変異によって引き起こされます。ミトコンドリア病は、ミトコンドリアDNAの変異や核DNAの変異によって引き起こされますが、多くの場合、家族性の遺伝ではなく、突然変異によって現れます。
ミトコンドリアの機能が低下すると、細胞の活動が低下し、次のような症状が現れます。
脳の神経細胞の場合は、見たり、聞いたり、物事を理解したりすることが障害される
心臓の細胞の場合は、血液を全身に送ることがしづらくなる
筋肉の細胞の場合は、運動が障害されたり、疲れやすくなったりする
ミトコンドリアの機能を活性化させるには、次のような方法があります。
汗ばむような運動を続けることで、ミトコンドリアの増加と活性が促される
バランスのいい食事を摂る
コエンザイムQ10を含む食品を摂取する
ビタミンB、マグネシウム、鉄などの栄養素を摂取する
癌細胞にもなりそうな異常な細胞を自殺させるアポトーシスとは何でしょうか?
アポトーシス(apoptosis)とは、細胞がミトコンドリアが自然死させる現象で、個体の成長や周辺のさいぼうに迷惑をかけずにきれいに死んでもらうために生命維持において重要な役割を果たしています。細胞自体に組み込まれた遺伝子によってプログラムされた、あらかじめ予定されている細胞の死で、細胞外からの障害によって死ぬ「ネクローシス」(壊死)と対義語として用いられます。しかし癌は活性酸素や放射能や紫外線がDNAを損傷させて遺伝子が突然変異させて起こるのではないのです。DNAの損傷と遺伝子が突然変異とは全く別のものなのです。ところが一方ヘルペスは細胞に感染すると細胞のゲノムにも必ず感染してしまうのでヘルペスウイルスは日常茶飯事的に遺伝子を変異させているのです。ヘルペスが多く感染している人の細胞は2分裂する度にヘルペスは何十倍、何百倍と増えていくので細胞のゲノムに感染する数が圧倒的に多くなりその分、2万3500個の遺伝子の中に含まれている800個あまりの細胞増殖関連遺伝子である増殖遺伝子と増殖抑制遺伝子を変異させる確率がどんどん増えていくのです。その結果、その確率が実現してしまうと二種類の増殖遺伝子と増殖抑制遺伝子が変異してしまうと異常なタンパクが生まれてしまい細胞分裂する必要がないのに変異増殖遺伝子は「増える」機能を獲得してしまい、他方変異増殖抑制遺伝子は「増えさせない」機能を失ってしまいその相互作用のために不必要な細胞が正常である細胞の遺伝子であるときよりも増え続けるのが増殖過剰細胞に過ぎないのです。つまり150年前から医療界が喧伝しまくる怖い怖いわけのわからない「癌細胞」などは存在しないのです。つまりヘルペスが突然ではなく普通に何の変哲でもなく細胞に感染し、さらにゲノムの中に感染し塩基の並びを変異させてしまった遺伝子が偶々、正常な細胞の増殖遺伝子と正常な細胞の増殖抑制遺伝子であれば人体の40兆個の細胞に一個のいわゆる癌細胞が誕生するだけであり突然に増殖遺伝子と増殖抑制遺伝子が突然変異が起こるわけではないのです。
「癌」という病名が持つ「死ぬ」という間違ったイメージが「癌は死ぬ病気である」と思い込まされて絶対に直すことができない三大標準医療である①手術②放射能③抗がん剤を実行投与して資本主義の目的であるお金を儲けるために治る癌を直せなくして故近藤誠さんのいう「早死に」をもたらしているだけなのです。②放射能③抗がん剤の二つは正に癌を起こしてしまう原因を作っているに過ぎないのです。さらに①の手術はいわゆる癌細胞の原因であるヘルペスが大量に巣くっているヘルペス感染細胞塊を除去しているだけで特別な癌病巣を除去しているわけではないのです。しかしherpesが大量に感染している塊を一時的に除去するだけで人体に感染しているヘルペスをしかも摘除するヘルペス感染細胞塊が大きければ大きいほど人間にとって絶対に必要な組織の欠如による人体に対する副作用がいかに巨大出るかを医者は金儲けのために説明しないのです。故近藤誠さんのいう「早死に」はまさに癌の手術なのです。何故ならば「手術」をやってしまうと元の組織は死ぬまで戻らないからです。②放射能③抗がん剤はいつでも辞められるうえに②放射能③抗がん剤治療では組織はなくなることがないからです。
アポトーシスが起こると、細胞は次のような変化をします。萎縮して隣接細胞から離脱します。クロマチンの凝縮、核の断片化、 細胞膜が壊れないで細胞の断片化。
細胞の内容物が漏れ出す前に、周辺の細胞がアポトーシス細胞を貪食するため、炎症を伴いません。ここで癌細胞もアポトーシスで殺すことができる最高のサイトカインであるインターフェロンα(IFN-α)について詳しく述べておきましょう。
インターフェロンα(IFN-α)とは、体内で生成されるタンパク質で、抗ウイルスや免疫調節、細胞増殖抑制作用などの働きがあります。感染症やガン、ヘルペスウイルスが起こすあらゆる自己免疫疾患の治療薬としてさらにあらゆる癌を引き起こすので慢性骨髄性白血病やヘアリー細胞白血病、多発性骨髄腫などの治療に用いられています。
インターフェロンαの主な働きには、①アポトーシス(計画された自殺)を誘導する②herpesが感染している標的細胞に抗ウイルス活性や免疫調節活性を持つ③異常な細胞増殖を阻害する④腫瘍細胞に対する生体の応答力を高める⑤NK細胞やマクロファージを活性化して腫瘍細胞に対する細胞障害性を高める。このようにインターフェロンα(IFN-α)は殺し切れない従ってワクチンが作れないヘルペスウイルスが感染した細胞をアポトーシスさせるために特化した進化の中で生まれたinterferon(インターフェロン)なのです。なのに何故ヘルペスが生み出した癌細胞を自殺に追い込むことや増えないように抑制することができないのでしょうか?細胞を自殺させる以上に ヘルペスウイルスが増えすぎてしまうからなのです。
インターフェロンには、α、β、γの3種類がありますが、αとβは似た性格をしており、γはα、βに比べて免疫調節作用や抗腫瘍効果、種特異性などが異なります
インターフェロンβ(IFN-β)は、線維芽細胞IFNとも呼ばれ、Ⅰ型インターフェロンファミリーに属する分泌型タンパク質です。主に線維芽細胞で産生されますが、病原体と出会った樹状細胞やマクロファージや、血管の内皮細胞も産生します。インターフェロンβの主な作用は、①腫瘍細胞表面に結合して増殖を抑制することと、②宿主を介して抗腫瘍免疫能を活性化することです。また、ヘルペスによる自己免疫疾患である多発性硬化症(MS)の進行を遅らせる治療薬としても使用されているのは当然なのです。インターフェロンα、β
は癌細胞を作ったヘルペスウイルスが最も恐れる免疫の遺伝子が製造した最高の「特効薬」なのですがこの「特効薬」の製造能力よりもヘルペスウイルスの増殖力がはるかに優れているのでた免疫が追い付かないからです。
アポトーシスは、生物の発生における細胞死や異常な細胞の除去を行う機構で、オタマジャクシがカエルになるときに尻尾が消失したり、脊椎動物の指の間の水掻きが胚の発生にともない、不必要な器官を除去する役割も果たしているのです。
アポトーシス(apoptosis)による細胞の死は体の中で実はいつも起こっているのです。組織が損傷されて引き起こされるネクローシスと呼ばれる細胞の壊死は近年では交通事故多いのですが日常的に起こる細胞死の大半はこのアポトーシス(apoptosis)によるものなのです。日常的に起こる細胞死の大半の大部分はこのインターフェロンαが関わっているのです。何故ならばストレスが極めて多い現代ではヘルペスウイルスが増えるばかりですからヘルペス感染細胞がうなぎ上りに増え続けているからです。最後はヘルペスが起こす癌で死ぬのです。近頃は3人の男性が癌になり2人が癌死します。インターフェロンαによるアポトーシス(apoptosis)の最大の特徴はヘルペスが周辺の細胞に感染しないように「自己犠牲の精神」をもって無限に増えていくヘルペスを自分の細胞もろとも迷惑をかけずに美しく自死することです。つまり細胞膜が破れる前にマクロファージのような食細胞(貪食細胞)が処理してくれるように「核」や「細胞骨格」重要な細胞成分が細胞の酵素で分解されてしまうのです。マクロファージが自分の細胞を食べてくれるのはアポトーシスを起こす細胞膜では普段は脂質二重層の内側に隠れている「ホスファチジルセリン」というリン脂質が「反転」して外側に露出しているホスファチジルセリンを食細胞が見つけてくれるからです。
herpesウイルスと一緒に自殺してくれる細胞のアトポーシスは遺伝子的にもきちんと制御された一連の反応であり二つの誘因があります。
一つ目は細胞外からアポトーシスを促進する刺激が与えられたときです。このような刺激はFas受容体からくるか、核のDNA損傷から来ます。
二つ目はアポトーシスを抑制している生存因子がなくなったときです。例えば発生中の神経細胞は成長因子である神経成長因子(nerve growth factor;NGF)がアポトーシスを抑制しているが生存因子である神経成長因子が欠如してしまうと計画死である細胞死(アポトーシス)に至るのです。逆に言えば発生中のがん細胞では生存因子である様々な増殖関連因子が正常な増殖には必要ないのに計画死である細胞死(アポトーシス)が出来なくなって癌細胞は増え続けるのです。言い換えると癌細胞外からアポトーシスを促進する細胞表面のFas受容体からも核のDNA損傷からも来ているにもかかわらず莫大な量の生存因子からの
増殖の刺激が来ているので癌細胞は増え続けるのです。この刺激をもたらし続けるのが細胞に感染したヘルペスが正常な増殖関連因子の二つである増殖因子と増殖抑制因子の両方の遺伝子を変異(いわゆる突然変異)させてしまったからです。
Fas受容体とは何ですか?アポトーシスは、生体の恒常性維持(ホメヲスターシス)に重要であり、生命維持に欠かせない細胞活動の一つです。Fasは免疫系で自分に強く反応する細胞をアポトーシスで除去する役割を持ち、がん細胞やヘルペスウイルス感染細胞の除去にも関わっています。Fasが働かないと、ヘルペス性の自己免疫性の関節リウマチや腎臓炎などの全身性の自己免疫疾患を発症します。Fasリガンドは、受容体であるFasに結合して細胞にアポトーシスを誘導する「死因子」となるサイトカインです。アポトーシスは、遺伝的にプログラムされた細胞死で、ほかの細胞にも一番怖いヘルペスを細胞もろとも殺すことをしなければならない生理的に自然に起こる細胞死なのです。
アポトーシスは、FasリガンドなどがFas受容体と結合するとFas受容体が活性化することによって誘導されます。FasリガンドとFas受容体との相互作用により、カスパーゼ8が活性化され、その後カスパーゼ8がシグナルを送り、下流のエフェクターカスパーゼを介してアポトーシスが開始され最後は死細胞になります。
核のDNA修復とは何でしょうか?核のDNAの修復とは、細胞内にあるDNAが損傷した際に、修復酵素によって傷を修復するメカニズムです。DNAはヒトの細胞の核内の染色体にあり、遺伝情報を伝える設計図として機能しています。DNAは、タバコなどの化学物質や紫外線などの環境因子、ヘルペス様なゲノムに組み込まれた内在性因子によって損傷を受けることがあります。DNAが損傷すると、タンパク質の機能不全や正常に機能しないタンパク質が生じ、早期老化や変性疾患、腫瘍形成につながる可能性があります。
DNAの修復には、①直接修復、②塩基除去修復、③ヌクレオチド除去修復(nucleotide excision repair;NER)などの機構があります。DNAの修復は細胞周期の4つの段階のいくつかの段階で起こりますが、細胞は主にG1/S期とG2/M期に停止してしまい細胞の遺伝子の複製も細胞分裂も停止してしまいます。さらにゲノム複製エラーにより損傷が生じた場合は、S期や有糸分裂後期であっても細胞周期は停止します。細胞周期とは細胞周期とは、細胞が分裂して2つの娘細胞を生み出すまでの過程を指します。
細胞周期とは何か?細胞周期とは増殖細胞が増殖刺激を受けると、「DNA複製」により細胞内のDNA量が2倍になり、「細胞分裂」の過程で均等に染色体が分配され、2つの娘細胞を形成します。この現象は周期的に起こることから「細胞周期」と呼ばれ、G1期→S期→G2期→M期の4つのステージを1方向のサイクルで進んでいます。
この細胞周期の進行を担っているものがCdk(Cyclin-dependent kinase; CDKで日本語訳はサイクリン依存性キナーゼ)と呼ばれるリン酸化酵素タンパク質で、周期を通じて安定的に存在をしているのですが、リン酸化活性は周期の進行に伴い変化します。このCdkはCdkとか大文字でCDK(Cyclin-Dependent Kinase)と書く場合もしばしば見られます。このリン酸化活性の制御をサイクリンというタンパク質が行っており、サイクリンがCdkに結合することでリン酸化能が活性化され、周期を進行させています。
細胞周期が進行する上で細胞外からのストレス(紫外線・発ガン剤など)によりDNAが損傷してしまうと、一旦DNAを修復するためG1期で停止します。これをG1チェックポイントといい、細胞周期のブレーキ役であるCdk阻害因子(cyclin-dependent kinase inhibitor;CKI)というタンパク質がCdkもしくはCdkーサイクリン複合体に結合し細胞周期の停止に関わっています。細胞周期の異常により起こる疾患の代表的なものにガンがありますが、ガン細胞の多くはG1チェックポイントの異常が大きく関わっています。
細胞周期をわかりやすくたとえて言うと「細胞」を国道環状上八号線を走る乗り物、「Cdk」はエンジン、「サイクリン」はアクセルで「CKI」をブレーキと表すことができます。もし、この国道環状上八号線で事故が発生したとすると、善良なドライバーは危険を察知してブレーキを踏み停止をしますが、暴走族のような悪いドライバーは停止せずに走り続けます。この悪いドライバーが「細胞周期」における「細胞ガン」なのです。
このように増殖細胞では、上記のタンパク質をはじめ、異常を察知するタンパク質など様々なタンパク質同士が相互作用をすることで、一連の周期制御(DNA複製・細胞分裂・ガン化抑制・・・など)を行うためのネットワークを形成し、外部からのシグナルに対して速やかに応答することができるのです。
細胞周期とは、G1期、S期、G2期、M期の4つの段階から構成されており、細胞の新たなる成長やDNAの複製、細胞分裂などのイベントが含まれています。細胞周期の「周期」という意味は「一回り」という意味で「細胞周期」の意味は「新旧の細胞を入れ替える時に4つの段階を一回りすれば」新しい細胞が生まれますよ」という意味になります。G1期、S期、G2期、M期の4つの段階は、次のような特徴を持っています。
細胞の成長と分裂は一連の過程を繰り返して行われており、その過程はいくつかの段階に分けられる。細胞周期は、 各期からその次の期へ進むかどうかを監視しているサイクリンとサイクリン依存性キナーゼによって制御されているのです。
サイクリンとサイクリン依存性キナーゼが結合したものが「サイクリンーサイクリン依存性キナーゼ複合体」なのです。つまり①「サイクリン」と②「サイクリン依存性キナーゼ」と③「サイクリンーサイクリン依存性キナーゼ複合体」の三つはそれぞれ別のものなのです。
「サイクリン」は細胞周期特異的なサイクリンは、CDKと結合することでCDKのタンパク質セリン/スレオニンキナーゼ活性が活性化させます。
「サイクリン依存性キナーゼ」は細胞周期を司る核内キナーゼ群でサイクリンと結合してはじめて活性をもつ.CDK1(別名cdc2),CDK 2,CDK 4,CDK 6があり,細胞周期の異なった時点で活性化されている。活性は,CDK自身のチロシン,スレオニン残基のリン酸化と脱リン酸化で制御されている。
cdc2とはCDC2とも書き、サイクリン依存性キナーゼ1(CDK1)の別名で、細胞周期に関与するプロテインキナーゼです。真核細胞の細胞周期の制御に重要な役割を果たしており、特にG2/M期移行に必須とされています。細胞周期とは、細胞が分裂するための準備が行われ、M期で細胞が分裂を開始する一連の流れです。細胞周期が正しく進行しているか監視し、異常があった場合、その進行を一時停止したり、抑制したりする機構を細胞周期チェックポイント機構といいます。
細胞周期チェックポイント機構とは細胞周期が正しく進行しているかどうかを監視し、異常があった場合は細胞周期の進行を一時停止したり抑制したりする機構です。
細胞周期は、G1期、S期、G2期、M期の4つの期(Phase)に分類されており、各期が秩序正しく進行するように制御されています。細胞周期チェックポイント機構は、この各期が正しく進行しているかどうかを監視し、異常があった場合は進行を一時停止したり抑制したりする役割を担っています
「サイクリンーサイクリン依存性キナーゼ複合体」はサイクリンとサイクリン依存性キナーゼ(Cdk)の複合体は、細胞周期の進行に不可欠なタンパク質複合体です。ヒトでは、11種類のサイクリンと4種類のCdkが知られており、さまざまな組み合わせで複合体を形成して細胞周期を様々に制御しています。
サイクリンとCdkの複合体の働きには①サイクリンの濃度が上昇すると、Cdkと結合してキナーゼが活性化されます。②DNA合成や細胞分裂など特定の段階で必要なタンパク質をリン酸化します。
サイクリンとCdkの複合体と関連する因子には、①Cdk1とCyclin Bの複合体は、細胞周期をG2期からM期に移行させます。②Cdk2とCyclin Eの複合体は、細胞周期をG1期からS期に移行させます。③サイクリンDとCDK4/CDK6の複合体は、G1期からS期への移行に重要な因子です。④Cdkはがん治療の標的にもなっています。CDKを阻害すると腫瘍細胞の異常な成長を止めることができるため、CDK4とCDK6に注目した癌の研究が多く行われています。
サイクリン依存性キナーゼとはサイクリンとサイクリン依存性キナーゼCDKと結合してキナーゼが活性化され、特定の段階で必要なタンパク質をリン酸化します。サイクリンというタンパク質に捕捉された基質となるタンパク質がCDKという酵素と結合し、活性化されたサイクリン-CDK複合体のキナーゼ活性によって基質をリン酸化することです。
- G1期(G1 phase): ギャップ1期とも呼ばれ、DNA合成のための準備期間です。細胞は増殖因子の有無や栄養状態などを識別し、次の細胞周期のS期に入るか否かを決定します。必要となるすべてのタンパク質、リボソーム、およびその他の機構が合成される。そして十分な量が用意できるとS期(S phase)に移行し、ここでDNAが複製される。
- S期: DNA合成期です。DNAが複製されます。
- G2期(G2 phase): ギャップ2期とも呼ばれ、次のM期のための準備期間であり成長期です。
- M期(M phase): 分裂期です。初めて染色体が現れて有糸分裂が起こります。
細胞周期の各段階の開始と終結は、「サイクリン-サイクリン依存性キナーゼ(CDK)複合体」の活性状態によって規定されています。4段階の細胞周期を一回りする進行は、すべてサイクリン-サイクリン依存性キナーゼ(CDK)複合体のタンパク質の活性状態によって厳密に制御されてチェックされています。
「サイクリン-サイクリン依存性キナーゼ(CDK)複合体」の仕事は具体的にはなんでしょうか?サイクリンの濃度が上昇すると、CDKと結合してキナーゼが活性化され、特定の段階で必要なタンパク質をリン酸化します。サイクリンというタンパク質に捕捉された基質となるタンパク質がCDKという酵素と結合し、活性化されたサイクリン-CDK複合体のキナーゼ活性によって基質をリン酸化することです。サイクリン依存性キナーゼ(Cyclin-dependent kinase; Cdk)とサイクリン(Cyclin)の複合体は、細胞周期の進行を制御するマスターレギュレーターなのです。
細胞周期チェックポイント機構とは、細胞周期が正しく進行しているかどうかを監視し、異常があった場合は細胞周期の進行を一時停止したり抑制したりする機構です。細胞周期は、G1期、S期、G2期、M期の4つの期(Phase)に分類されており、各期が秩序正しく進行するように制御されています。細胞周期チェックポイント機構は、この各期が正しく進行しているかどうかを監視し、異常があった場合は進行を一時停止したり抑制したりする役割を担っています。
サイクリンとCdkの複合体の役割はサイクリン依存性キナーゼ(Cdk)とサイクリン(Cyclin)の複合体は、細胞周期の進行を制御する役割を担っています。細胞周期の進行にはこの複合体の存在が不可欠で、細胞周期の主要な調節因子として知られています。ヒトでは、細胞周期に関与する11種類のサイクリンと4種類のCdkが知られており、これらがさまざまな組み合わせで複合体を形成します。なかでも、Cdk1-サイクリンB複合体は分裂期への進行に必須の役割を果たしています。
さらに細胞周期の進行を制御する具体的なサイクリンとCdkの複合体(サイクリンーCdk複合体)の組み合わせとそれぞれの組み合わされたサイクリンーCdk複合体役割は何でしょうか?
- CDK1-サイクリンB複合体は分裂期への進行に必須の役割を果たす
- CDK4は細胞周期中のG1期からS期への移行に必須
- CDK inhibitor(CKI)はcyclin-CDK複合体に結合することでCDKのキナーゼ活性を阻害し、細胞周期の制御に重要な役割を果たす
ヒトでは細胞周期に関与する11種類のサイクリンと4種類のCdkが知られており、それらがさまざまな組み合わせで複合体を形成します。また、それぞれのがんやアポトーシスへの関与も異なります。
細胞周期は、ケガや病気によって細胞が死んでしまった場合に失われた細胞を補うために更に新旧の細胞を入れ替えるために日常的に繰り返し繰り返し行われています。サイクリンーサイクリン依存性キナーゼ(CDK)複合体は、サイクリンというタンパク質とサイクリンキナーゼという酵素蛋白の二つの蛋白の複合体です。サイクリンーサイクリン依存性キナーゼというのはサイクリンに依存して働くことができるキナーゼ(リン酸化酵素)の複合体であり細胞周期の進行を制御するタンパク質複合体となるのです。また、細胞周期の制御には、CDK inhibitor(CKI)と呼ばれるたんぱく質も重要な役割を果たしています。CKIはサイクリン-CDK複合体に結合することでCDKのキナーゼ活性を阻害します。
サイクリンー依存性キナーゼ(Cyclin-dependent kinase; CDK)とは、4つの段階を持つ細胞周期の主要な調節因子である酵素で、セリン・キナーゼとスレオニン・キナーゼの二つに分類されます。一方、サイクリンは細胞周期の進行をつかさどる酵素群のサブユニットを構成するタンパク質です。サブユニット(subunit)とは、次のような意味があります。
- 部品や部分構成要素を意味するビジネス用語
- 複数のタンパク質によって構成されるタンパク質複合体の構成単位となる単一のタンパク質分子でポリペプチド鎖と呼ばれます。
タンパク質の構造において、複数のポリペプチド鎖(サブユニット)が集合して形成したものを四次構造といいます。この四次構造を形成する各ペプチド鎖がサブユニットと呼ばれます。また、遺伝子の翻訳を行い蛋白を作るリボソームという巨大なRNA・タンパク複合体も、大小2つの粒子に分かれており、それぞれ50Sサブユニットと30Sサブユニットと呼ばれています。
サイクリンとCDK (Cyclin-dependent kinase)は、サイクリンもCDKも多種類があるのでさまざまな組み合わせで複合体を形成し、4つの段階の細胞周期の各段階の進行を厳しく制御し「進め」「止まれ」の信号を出します。サイクリンに捕捉された基質となるタンパク質がCDKと結合し、サイクリン-CDK複合体のキナーゼ活性によってリン酸化を受けます。
ヒトでは、細胞周期に関与する11種類のサイクリンと4種類のCDKが知られており、それぞれのがんやアポトーシスへの関与も異なります。
複製エラーとは、DNAの複製時にDNAを構成する塩基を間違えて写し取ることです。鋳型と異なる塩基を複製したり、あるはずの塩基を複製しなかったり、余分な塩基を複製したりする場合などが該当します。
DNA複製エラーとは、DNAポリメラーゼというDNA複製酵素が10万回に1回という高頻度で引き起こします。単純計算すると、1回のDNA複製で6万文字も誤って複製されてしまうことになります。この複製エラーとは、DNAの複製時にDNAを構成する塩基を間違えて写し取ることです。鋳型と異なる塩基を複製したり、あるはずの塩基を複製しなかったり、余分な塩基を複製したりする場合などが該当します。
複製エラーは遺伝子の突然変異を引き起こし、細胞のがん化や遺伝病の原因ともなりますが生物はミスマッチ修復と呼ばれる複製の間違いを修正するための防御システムを持ち、複製エラーを防いでいます。ミスマッチ修復(MMR)とは、DNAの複製中に発生するミスマッチ塩基対を修復するシステムです。ミスマッチ塩基対とはDNAポリメラーゼが誤った残基を取り込むことでミスマッチ塩基が生じますが、ミスマッチ修復によって、このミスマッチ塩基を修復し、複製の忠実度を高めています。DNAは4つの塩基であるGに対してCと、Aに対してTの互いに相補的な水素結合基を有する核酸塩基が水素結合した塩基対から形成されている。これらG―C、A―T塩基対をWatson-Crick塩基対と呼ぶ。一方、これらG―C、A―T以外にも核酸塩基の組み合わせはG―G、G―A、G―T、A―A、A―C、C―T、C―C、T―Tの8種類可能である。これら8種類の塩基対になるとミスマッチ塩基対と呼ぶ。ミスマッチ塩基対はWatson-Crick塩基対に比べると、熱力学的な安定性が低下しミスマッチ塩基対を含む二重鎖は、完全に相補的なDNA二重鎖と比べて低い温度で二重鎖の解離が起こる。ミスマッチ塩基対はDNA中ではDNAポリメラーゼが誤った残基を取り込む複製エラーにより生じるが、修復酵素により除去される。バルジ構造と同じくRNAにはごく普通にみられる構造である。バルジ構造とは二重鎖核酸の塩基対の一部が塩基対合できず、飛び出た部分です。バルジの意味は「飛び出るとか膨らむ」の意味があります。しかもゲノムの1.2%しかない23500個の遺伝子のミスマッチ塩基対の修復は誤りなく行われるといわれます。ミスマッチ塩基対とは残りの98.8%のミスマッチ塩基対の誤りは放置しても遺伝子には関りがないので何の問題も起こりません。
複製エラーは遺伝子の突然変異を引き起こし、細胞のがん化や遺伝病の原因ともなることがあるのはヘルペスが起こした遺伝子の複製エラーは突然変異だけです。生物はミスマッチ修復と呼ばれる遺伝子の複製の間違いを起こさないほとんど完全な防御システムと遺伝子の複製の間違いを修復するシステムを持ち、複製エラーを防ぐことができるのです。
ミスマッチ塩基対が何故重大問題となるのですか? DNA 複製の際に DNA ポリメラーゼが誤った塩基を取り込むことで発生する、対合する塩基を持たないとか、正しい対にならないアンペア塩基とか通常とは異なる塩基同士が対合したミスペア塩基などの総称です。DNA は、A(アデニン)、T(チミン)、G(グアニン)、C(シトシン)の 4 種類の塩基から構成されており、A と T、G と C が結合して塩基対を形成しています。この塩基対は、DNA の構造を明らかにしたワトソンとクリックにちなんで「ワトソンクリック塩基対」とも呼ばれます。
ミスマッチ塩基対は、DNA 鎖の 1 本に誤った塩基が組み込まれることで発生する複製エラーによって形成されます。次の複製サイクルでも、この鎖が DNA 合成のテンプレートとなり新しい塩基対を形成し続けるので後々まで新たなるミスマッチ塩基対が生まれ続けるからです。
さてアポトーシスの分子生物学的な意味と癌細胞とヘルペスウイルスとの関りは何でしょうか?
アポトーシス細胞の内部構造を維持しているタンパク質である核ラミンや細胞骨格のタンパク質などの分解が細胞のあちこちで一斉に生じるという特徴がある。核ラミンとは、核膜の内膜の裏側にある網目状の繊維構造で、ラミンというタンパク質が重合してできています。核の形態を支え、核膜とクロマチンの相互作用を調節することで、核内のさまざまな反応に関与しています。核ラミンは、細胞骨格の1つである中間径フィラメントを構成するタンパク質で、DNA複製や遺伝子発現、クロマチン構造の制御などの細胞内プロセスにおいて重要な役割を担っています。ラミンは、配列相同性、発現パターン、生化学的な特徴からA型とB型のラミンに分類され、脊椎動物ではLamin-AとLamin-C、Lamin-B1とLamin-B2などが主要なラミンです。ラミンとその関連タンパク質に異常が生じると、疾患の原因となります。たとえば、ラミンA(Lamin-A)をコードするLMNA遺伝子に突然変異が生じると早老症を引き起こします。また、ラミンAやラミンCに関連する疾患には、筋ジストロフィーやシャルコー・マリー・トゥース病などの筋肉や神経系疾患があります。
核ラミンや細胞骨格のタンパク質などの分解の主役はカスパーゼ(caspase)と言われる蛋白分解酵素です。カスパーゼ(caspase)の名前は酵素の活性部位にシスティンがあり標的蛋白のアスパラギン酸のところを切断することから生まれたのです。つまりcystein aspartate proteaseの単語の一部である「c」と「as」と「ase」をとってきてcaspaseとつけたのです。カスパーゼ(caspase)は普段は活性のないプロカスパーゼの状態にあるのですがこのプロカスパーゼが切断されると活性化する性質を持っているのです。活性化したカスパーゼは別のカスパーゼを切断してさらにこのカスパーゼがまた別のカスパーゼを切断していくという連鎖反応が起こり最終的には細胞全体で分解反応が起こってしまうのです。
このようにカスパーゼは細胞内のシステインプロテアーゼの一種で、ミトコンドリアは細胞内エネルギーの生成や細胞死(アポトーシス)に重要な役割を果たす細胞小器官です。カスパーゼの活性化とミトコンドリアは、ともに細胞の生と死の両方の制御に関連しています。
カスパーゼとは、もっと詳しく説明すると蛋白質の切断を介してアポトーシスによる細胞死を実行する酵素です。アポトーシスは、個体をより良い状態に保つために異常な細胞が自殺できる「遺伝子にプログラムされた細胞死」で、厳密にコントロールされています。カスパーゼ-8はアポトーシスの開始に重要で、活性型カスパーゼ-8がカスパーゼ-3を切断することで細胞死が誘発されるのです。カスパーゼ-8およびカスパーゼ-3は、アポトーシス反応の重要な調節因子です。カスパーゼ-8はアポトーシスの開始に重要です。活性型カスパーゼ-8はカスパーゼ-3を切断し始め、その結果細胞死が誘発されます。カスパーゼ-3は、アポトーシスの重要なメディエーターの1つとしても同定されました。Bcl-2とBcl-XLを切断して、これらのタンパク質の抗アポトーシス機能を失わせると共に、アポトーシスを促進するC末端断片を遊離させます。アポトーシス活性化に後天的欠陥があると、多くの場合、細胞増殖の制御不良、発癌、および癌につながります。発癌と癌の違いは何でしょうか?発がんが正常な細胞の遺伝子が傷つくことで起こる現象であるのに対し、癌は正常な細胞の遺伝子が傷ついてできた異常な細胞が無秩序に増殖して発生する病気です。がんは、正式には「悪性腫瘍」と呼ばれ、「悪性新生物」とも呼ばれます。また、臓器名や組織名の語尾に「癌」という用語がつく場合は「癌腫」を意味します。癌腫とは、体の表面や消化管および気道などの内側、臓器などを覆う「上皮細胞」に発生する癌のことです。従って肺がんや胃がん、大腸がん、乳がん、子宮がんなどは癌腫のひとつに数えられます
腫瘍壊死因子(TNF)受容体にリガンドが結合すると、FADD(Fas-associated protein with death domain)や他のデスドメインアダプタータンパク質が動員され、それらがカスパーゼを動員・活性化することによってアポトーシスが開始されます。アポトーシス時、一連のカスパーゼ(カスパーゼ-2、-8、-9、-10、-11、および-12)が、下流における別の一群のエフェクターカスパーゼ(カスパーゼ-3、-6 、および-7)を活性化し、これが他のタンパク質を切断してアポトーシスのプログラムが完了します。一方、ミトコンドリアは細胞内エネルギーの生成に重要な役割を果たしており、細胞死(アポトーシス)でも中心的な役割を担っています。
また、カスパーゼは非細胞死性に制御する生理機能も明らかになってきており、細胞移動や細胞の運命決定、精子の成熟や神経軸索の刈り込みなどに関与しています。
哺乳類のアポトーシスにおけるカスパーゼ活性化経路とはなにか?Fasや腫瘍壊死因子(tumor necrosis factor: TNF)レセプターなどによる刺激が引き金になります。Fasなどのリガンドがレセプターに結合すると、アポトーシス促進性タンパク質Bax、Bad、Bid、Bak、Bimが活性化します。これらのタンパク質にはBH3デスドメインがあり、アポトーシス阻害タンパク質Bcl-2やBcl-XLの機能を低下させることで、アポトーシスシグナルがミトコンドリアに伝達されます。ミトコンドリアからはシトクロムcが放出され、Apaf-1、Caspase-9と複合体(Apoptosome)を形成し、カスパーゼカスケードが開始されてアポトーシスが起こります。
ミトコンドリア内のエネルギーを産生するために酸素を消費する過程である電子伝達系では、多量のエネルギー(ATP)が産生されると同時に、多くの疾患の発症や老化の進行と関わりがある活性酸素が常に発生しているといわれますが、これは嘘です。しかし活性酸素は老廃物であるので、このようにして生じた活性酸素がどのようにして除去されているのかを解説しますが活性酸素が癌をはじめとする諸悪の病気の根源あるように言われていますがミトコンドリア間違いであることをも証明して行きます。がんも怖い病気でないように活性酸素も恐れることもないのです。
1.活性酸素とは活性酸素の種類(狭義)。活性酸素とは、酸素分子よりも酸化力が強い酸素や酸素の誘導体のことで、一般的に「スーパーオキシド」「過酸化水素」「ヒドロキシラジカル」「一重項酸素」の4種類があります。ここでいう酸素分子とは、三重項酸素のことを指しています。物質の最も安定な状態は基底状態と呼ばれ、空気中にある酸素の大部分はこの基底状態の酸素(三重項酸素)になっています。酸素は基底状態が三重項で、励起されると一重項になる例外的な分子で、生体や食品を構成する物質の大部分は一重項状態が基底状態となっています。

三重項と一重項ってどういう意味?三重項状態とは同じスピンの向きの電子を2つもつ分子のことで、一重項状態とはスピンの向きが正負同数である分子のことをいいます。言葉の説明だけでは少しわかりにくいので、三重項酸素と一重項酸素の違いを図として載せておきます。
三重項酸素と一重項酸素の違いは何ですか?

実は、三重項酸素も高いエネルギーをもっていますが、三重項分子と一重項分子は反応しないという性質があるので、常温では基底状態の酸素(三重項酸素)は激しい酸化反応を起こしにくくなっています。
しかし、基底状態にある酸素が何らかのエネルギーによって励起されると一重項酸素に変化して、活性酸素としてさまざまな物質(多くの物質は一重項状態が基底状態になっています)と反応を起こすようになります。
フリーラジカルとは、不対電子をもつ反応性が高い物質のことです。物質は共有結合によって2つの電子が対となっていますが、何らかのエネルギーを受けると共有結合が切れてフリーラジカルを生じます。
A : B → A・ + B・
ラジカルは反応性が高く不安定な物質なので、生体分子などに多くの傷害を与えます。活性酸素のうち、ヒドロキシラジカルやスーパーオキシドはフリーラジカルの一種であるのはこれらは不対電子をもっているからです。このうち、ヒドロキシラジカル(・OH)は活性酸素の中で最も反応性が高く、最も酸化力が強いフリーラジカルです。
ヒドロキシラジカルは過酸化水素への紫外線の照射や、過酸化水素と2価の鉄イオンの反応(フェントン反応)などによって生成されます。

スーパーオキシドアニオンラジカルとはスーパーオキシドアニオンラジカルはスーパーオキシドと呼ぶことが多いのですが、酸素が電子を1つ受け取って、アニオン(陰イオン)に変化したものです。スーパーオキシドはヒドロキシラジカルとは違い、それ自体の反応性はそれほど高くありません。しかし、スーパーオキシドからは種々の反応によって過酸化水素やヒドロキシラジカルなどの活性酸素を生成しますので、活性酸素として非常に重要な分子となっています。
2.ミトコンドリアでの活性酸素の発生と除去。ミトコンドリア内の電子伝達系では、酸素を消費する過程で、スーパーオキシドや過酸化水素といった活性酸素が発生しています。

この過程で生じるスーパーオキシドや過酸化水素は、それぞれSOD(スーパーオキシドジスムターゼ)やカタラーゼといった抗酸化酵素によって取り除かれています。
①スーパーオキシドジスムターゼとは何か?SOD(スーパーオキシドジスムターゼ)はその名の通り、スーパーオキシドを不均化する酵素のことで、2分子のスーパーオキシドを不均化して過酸化水素と酸素に分解します。ここでの不均化とは、2つの同一種の物質から異なる二種類の物質を生じる反応のことをいいます。

この反応で生じた過酸化水素は、さらにカタラーゼやグルタチオンペルオキシダーゼによる分解を受けて水に変換されます。
②カタラーゼとは何か?カタラーゼは、2分子の過酸化水素を不均化して水と酸素に変換させます。

③グルタチオンペルオキシダーゼとは何か?グルタチオンには還元型と酸化型の2種類があり、還元型グルタチオン(GSH)はその還元力を使い、活性酸素である過酸化水素(H2O2)を水(H2O)と酸素(O2)に変えることによって活性酸素の毒性を無毒化していますが、この反応はグルタチオンペルオキシダーゼという酵素によって行われています。ここでのペルオキシダーゼとは、酸素を2つ連続してもつ物質(ペルオキシ構造-O-O-をもつ物質)を酸化的に分解する酵素のことをいいます。還元型グルタチオンは基質を還元しますが、同時に還元型グルタチオン自身も基質によって酸化されるので、還元型グルタチオンは酸化型グルタチオン(GSSG)に変化してしまいます。生体ではペントースリン酸経路由来のNADPHの還元力を利用することで還元型グルタチオンを再生しています。「ペントースリン酸経路の役割」とは体内で消費されるグルコースのほとんどは,解糖系でピルビン酸に変換される過程でエネルギー産生に使用される。ペントースリン酸回路は,細胞に必要な産物である五炭糖(C5)とNADPHを得るために,グルコースが代謝される経路です。五炭糖のリボースあるいはデオキシリボースは核酸の構成成分として使用され,NADPHは脂肪酸合成の際に必要な補酵素なのです。
グルタチオンペルオキシダーゼによるH2O2の分解図

酸化型グルタチオンとは何でしょうか?酸化型グルタチオンは2分子のグルタチオンが-SH(チオール基)どうしで結合したものです。活性酸素の除去には、いわゆるスカベンジャーと呼ばれる、ビタミン類(CやEなど)やポリフェノールといった抗酸化物質によっても行われています。スカベンジャー(scavenger)とは、動物の死骸を食べる動物をスカベンジャーと呼び、死肉を食べる行為をスカベンジングといいカニ、カラスなどです。他に①動物の死骸やごみを食べる動物、またはその行為②化学反応の終了後に残存する特定の成分を捕捉して除去し処理する物質③体内の不要物質や毒性物質を処理できる器官・細胞・物質など④化学反応の終了後に残存する特定の成分を捕捉して除去する物質もスカベンジャーと呼ばれます。
グルタチオンは分子内にチオール基(-SH)を有する低分子化合物であり、これが酸化反応を受けるとジスルフィド(-S-S-)結合を介して二量化する。グルタチオン分子内にチオール基(-SH)を有する低分子化合物を還元型グルタチオン(GSH)、酸化反応を受けるとジスルフィド(-S-S-)結合を介して二量化したグルタチオンを酸化型グルタチオン(GSSG)と呼び、一般的に抗酸化作用やグルタチオン抱合を示すのは還元型なのです。
疲労を起こすのは活性酸素による酸化ストレスで、神経細胞が破壊されるからであると考えられていますが間違いです。運動などのエネルギーをたくさん使う活動では、酸素が多く消費されるとともに活性酸素も多量に発生します。活性酸素が発生すると、活性酸素を分解して体内から除去する抗酸化酵素が働くようになっていますが、発生する活性酸素の量が抗酸化酵素の働きを上回ると自律神経の細胞や筋肉が活性酸素によって攻撃されて疲労へとつながり休養すれば回復します。発生する活性酸素の量が抗酸化酵素の働きを上回ると自律神経の細胞や筋肉が活性酸素によって攻撃される根拠は何もないので仮に活性酸素がDNAを損傷しても素早くDNA修復機構が働いて即座に修復してくれます。かりに活性酸素の量が抗酸化酵素の働きを上回りすぎれば酸素を使てエネルギーとなるATPは作れませんからあらゆる活動は不可能になりますので疲労の限界を萌えることになります。睡眠や休養をいくらとっても疲労が解消されない時に疲労の他に、頭痛、のどの板城、筋肉痛、関節痛、睡眠障害、思考力低下や集中りぃくのていか、さらに美根湯、首のリンパ節の張れ、筋力低下などの症状が現れ強い疲労状態が6か月以上続いた時に「慢性疲労症候群」さらに「慢性疲労症候群」と診断されます。原因は長期にわたる心身への過剰なストレス係免疫を抑制しすぎてヘルペスが全身に増殖しすぎて特に多数のさまざまな症状が自律神経節に感染増殖したヘルペスと免疫との戦いが始まり炎症症状が出現し続けてしまうのです。
治療法はまずストレスの原因が何かを突き止め解消すべきです。次に抗ヘルペス剤と漢方のせんじ薬を大量に服用すれば必ず治ります。ヘルペスとの戦いによる炎症で痛みが出る機序はherpesウイルスと自然免疫細胞が戦うと発痛物質の代表であるであるブラジキニンが作られます。そと後に発痛物質の痛みの作用を強めるプロスタグランジンという生理活性物質も作られ痛みを増幅するのです。さらにストレスが長引けば長引くほど免疫が下がりherpesが増え続けると痛みを感知する全身のあちこちにある自由神経終末のレセプターに感染することによって激しい痛みも生まれるのです。
さらに「慢性疲労症候群」で発熱はどのようにして生まれだるさが生まれるのでしょうか?ストレスが「慢性疲労症候群」の原因はストレスでヘルペスウイルスが増えます。「慢性疲労症候群」で体温があがるのは、身体の中で免疫細胞がヘルペスウイルスと闘っている証拠です。体温を上げて免疫を活性化させ、ウイルスへの攻撃力を高めているのです。
発熱は、体に備わった体温を調節する仕組みによって起こります。通常は、私たちの身体は37℃前後に保たれています。これは、脳にある体温調節の司令塔である視床下部が設定した温度です。これをセットポイントといいます。ウイルスに感染すると、免疫細胞が察知して発熱物質を作り、ウイルスが体内に侵入したことを視床下部に伝えます。そして免疫を活性化させるために、体温を調整する機能が作用して、セットポイントを高い温度に設定します。そうすると、脳から身体に発熱するように指令が出されるのです。てプロスタグランジンが発熱指令となり視床下部に設定温度を新たに高温にセットさせなおすのです。痛みと発熱にエネルギーを注ぐために正常な体のきのうの訳20%は低下してしまいます。その結果、運動をし過ぎてエネルギーを使い過ぎた後の疲労の状態になってしまうのです。つまり痛みも発熱もブラジキニンからはじまりプロスタグランジンへと指令が伝わっていくのです。
熱が出る原理は?ウイルスに感染すると、免疫細胞が察知して発熱物質を作り、ウイルスが体内に侵入したことを視床下部に伝えます。 そして免疫を活性化させるために、体温を調整する機能が作用して、セットポイントを高い温度に設定します。 そうすると、脳から身体に発熱するように指令が出されるのです。
抗酸化作用を持つ物質には、①ビタミンC、②ビタミンE、③ビタミンA(レチノールやカロテンなど)④ポリフェノール(フラボノイド系や非フラボノイド系)④カロテノイド⑤コエンザイムQ⑥イオウ化合物⑦グルタチオン⑧アセチルシステイン
抗酸化物質が含まれている食べ物や飲み物には、次のようなものがあります。
アントシアニン(赤ワイン)
イソフラボン(大豆)
ケルセチン(玉ねぎ)
カテキン(緑茶)
テアフラビン(紅茶)
セサミン(ごま)
セサミノール(ごま)
クルクミン(ウコン茶、カレー粉)
ルチン(そば、柑橘類、玉ねぎ)
カカオポリフェノール(チョコレート、ココア)
また、抗酸化作用のあるサプリメントには、ビタミンA(βカロテン)、ビタミンC、ビタミンE、リポ酸、コエンザイムQ10などがあります。
抗酸化酵素には、スーパーオキシドディスムターゼ(SOD)、カタラーゼ、グルタチオンペルオキシダーゼ(GSH-Px)などがあります。
抗酸化作用とは、活性酸素の発生や働きを抑制したり、活性酸素そのものを取り除く作用です。活性酸素は病気や老化や癌の原因となるため、抗酸化作用を持つ物質を摂取することで、アンチ・エイジングや健康の維持に役立ちますといわれ、さらに活性酸素が病気や老化や癌の原因となるといわれますがその根拠が一切語られていないのです。つまり活性酸素はDNAの損傷をもたらしDNAの傷が蓄積して最後は遺伝子が癌化するといわれていますが大間違いです。一つ目の間違いはDNAが即遺伝子ではないのです。二つ目の間違いは遺伝子となっているDNAの損傷はDNA修復機構を形成している遺伝子がすべて修復することが100%証明されています。できるのです。三つ目はアンチ・エイジングは生理的老化を防ぐことですが無理です。何故ならば人間の老化には4原則があります。生理的老化の特徴を端的に表すものとして、「シュトレーラーの4原則」が知られています。
- 普遍性。生理的老化は、誰にでも普遍的に起こるもの。自分だけ不老不死を願って何をやっても、生理的老化を避けることはできない。
- 内在生。生理的老化は、遺伝的にプログラムされているもので生理的老化が進行しないよう環境を改善しても、限界があるからです。
- 進行性。生理的老化が進んだら、もとに戻ることはない。
- 有害性。ヒトにとって、生理的老化は誰にとっても有害であり、一般的に有益ではない。
4つ目の健康の維持は抗酸化作用を持つ物質を摂取することで活性酸素の発生や働きを抑制できるといわれますが健康の維持のために老人も運動もやりなさいと勧められますが運動をやるたびに活性酸素が増えるのですよ。おお大いなる矛盾ですね。若い時から運動を続けている人が酸素を使いすぎて活性酸素の産生が最も多いので癌も多いはずなのですがそんなデータは目にしたことがありません。
抗酸化作用のある物質は、食事や飲み物やサプリメントからから摂取できますと大いに宣伝されていますが医学でも処理不可能といわれている活性酸素を食事や飲み物やサプリメントで人体から除去できると思いますか?不可能です。
ビタミンCは水溶性なので、生で食べるのがよいとか、パプリカやブロッコリー、じゃがいも、キウイ、柑橘類、イチゴ、柿などに多く含まれていますといわれていますが好きなようにしてください。
コーヒーにはコーヒーポリフェノールという抗酸化作用を持つ成分が含まれています。β-カロテンはトマトやホウレンソウ、ピーマン、ブロッコリーなどの緑黄色野菜に多く含まれますとも耳にしますが適当にやってください。
グルタチオンには、次のような効果が期待できます。
解毒作用によって疲労やストレスを軽減する
アルコール摂取によって生じるアセトアルデヒドを効率よく分解する
肝臓や腎臓の働きを阻害するミネラルを体外に排泄する
飲酒のし過ぎによって生じた脂肪肝の改善に効果がある
肝障害の予防、改善につながる
グルタチオンは、年齢や紫外線、ストレス、飲酒、喫煙などの影響で減少していきます。体内のグルタチオン量を増やすには、「シスチン」と「テアニン」を合わせて補給するとよいでしょうとかまた、食品ではブタ肝臓、肉類(牛肉、豚肉)、パン酵母、牡蠣、アボガド、トマト、小麦粉、お米などに含まれていますとか言われますが誰もが普通に食べているものですね。アッハッハ!!
グルタチオンとは何でしょうか?
グルタミン酸、グリシン、システインという3つのアミノ酸が連なったペプチドで、人間の体内に広く分布する物質です。強力な抗酸化作用があり、身体を活性酸素のさびつきから守る極めて大切な役割以外に様々な作用を持っています。7つの主な作用を列記しましょう。
グルタチオン(GSH)の作用
1.抗酸化作用
2.解毒作用
3.免疫力強化
4.抗酸化脂質・抗動脈硬化
5.エネルギーレベルの復元・活性化
6.放射線損傷の保護と修復
7.ミトコンドリア合成の促進
8.虚血・再灌流障害
1.細胞を活性酸素から守る強力な抗酸化作用があります。
① 食品添加物、薬品(アセトアミノフェン等)、化学物質、アセトン(除光液など)、農薬、重金属(水銀、鉛、カドミウム、など)、人口甘味料、洗剤、消臭スプレー、カビ取りクリーナー、紫外線、放射線、電磁場、大気汚染物質、ショック、虚血、虚血再灌流、精神的ストレス、肉体的ストレス、偏った食事、アルコール、タバコ、酸化された食べ物、人口着色料、人口甘味料、炎症、過酸化物、過度な運動などによって過剰に発生した活性酸素から細胞を守る。② グルタチオンは細胞内で電子を失って不安定な状態になったフリーラジカルに電子を与えて安定分子に戻し、細胞膜や核酸などを傷つけないようにする。GPx(グルタチオンぺルオキシダーゼ)は細胞内(ミトコンドリア内)でSODによってスーパーオキシド(・O2-)が除去された後に残った過酸化水素(H2O2)を除去するのに触媒として働く。
グルタチオンには、次のような効果が期待できますといわれていますが
解毒作用によって疲労やストレスを軽減する
アルコール摂取によって生じるアセトアルデヒドを効率よく分解する
肝臓や腎臓の働きを阻害するミネラルを体外に排泄する
飲酒のし過ぎによって生じた脂肪肝の改善に効果がある
肝障害の予防、改善につながる
グルタチオンは、年齢や紫外線、ストレス、飲酒、喫煙などの影響で減少していきます。体内のグルタチオン量を増やすには、「シスチン」と「テアニン」を合わせて補給するとよいでしょう。
また、食品ではブタ肝臓、肉類(牛肉、豚肉)、パン酵母、牡蠣、アボガド、トマト、小麦粉、お米などに含まれています。
1.抗酸化作用(細胞を守る)①食品添加物、薬品(アセトアミノフェン等)、化学物質、アセトン(除光液など)、農薬、重金属(水銀、鉛、カドミウム、など)、人口甘味料、洗剤、消臭スプレー、カビ取りクリーナー、紫外線、放射線、電磁場、大気汚染物質、ショック、虚血、虚血再灌流、精神的ストレス、肉体的ストレス、偏った食事、アルコール、タバコ、酸化された食べ物、人口着色料、人口甘味料、炎症、過酸化物、過度な運動などによって過剰に発生した活性酸素から細胞を守る。②グルタチオンは細胞内で電子を失って不安定な状態になったフリーラジカルに電子を与えて安定分子に戻し、細胞膜や核酸などを傷つけないようにする。GPx(グルタチオンぺルオキシダーゼ)は細胞内(ミトコンドリア内)でSODによってスーパーオキシド(・O2-)が除去された後に残った過酸化水素(H2O2)を除去するのに触媒として働く2GSH+・O2-GPxGS-SG+2H2Oまた過酸化脂質も同様に還元・無毒化される。LOOH+2GSHGPxLOH+GS-SG+H2O(Lは脂質(Lipid))③グルタチオンは細胞内で98%以上が還元型として存在するが、酸化ストレスにさらされると酸化型を還元型に変換する酵素(グルタチオンレダクターゼ)が活性化され、酸化型グルタチオンは還元型グルタチオンにリサイクルされる。GS-SG+NADPH+H+2GSH+NADP+このリサイクルが素早く行われないと酸化型グルタチオンは細胞の外へ排出されてしまうため、フリーラジカルが大量に発生する状況では、リサイクルが間に合わなくなりグルタチオンは枯渇してしまう。還元型グルタチオンの濃度は一定に保たれているが、何らかの理由により、還元型と酸化型の比率が大きく崩れた場合は、酸化型グルタチオンが細胞外に積極的に排出される。ミトコンドリアとグルタチオンブドウ糖ATPATPミトコンドリア①ブドウ糖からATPを産生する時、ミトコンドリアは活性酸素を発生する。②細胞内のグルタチオンなどのスーパーオキシド・O2-フリーラジカル・スカベンジャーが活性酸素を捕まえて消去する。H2O2フェントン反応③しかし年齢と共にグルタチオンなどの産生が減少し、細胞はH2O+O2・OHミトコンドリアが発生した活性酸素ヒドロキシラジカルを処理できなくなる。DNA損傷④ミトコンドリアも細胞も損傷し、細胞膜損傷細胞死に追いやられる。SODGSHFe2.解毒作用①グルタチオンは主に肝細胞においてアルコール、薬物(抗生物質など)毒物、重金属、伝達物質(ロイコトリエン、プロスタグランジンなど)など異物を細胞外に排出する。②まず、有害物質をグルタチオンが包み込み、水に溶ける状態に変化させる。グルタチオンはグルタチオン-S-トランスフェラーゼ(GSTs)によってシステイン残基のチオール基に有害物質を結合する。これをグルタチオン抱合という。③グルタチオン抱合した有害物質は、グルタチオンと共に細胞の外へ押し出される。④その後腎臓まで運ばれた抱合体からグルタミン酸とグリシンが切り離され、残ったシステインと有害物質がメルカプツール酸となって胆汁中や尿中に排出される。⑤肝臓のグルタチオンが毒物や異物に結合して消費されると、γーグルタミルシステイン合成酵素が活性化され、グルタチオンが合成される。⑥有害物質の量が肝臓内に存在するグルタチオン以上に多い場合は、有害物質が処理できずに残り、肝細胞を傷つけることになる。⑦γーGTPは膜タンパク酵素であり、膜に結合して存在し細胞外のグルタチオンを分解する。グルタチオンはそのままでは、細胞内に入れないので、分解されたアミノ酸が細胞内に入って再合成されることになる。酸化ストレスが高く活性酸素が多量に出ている状態では、細胞内のグルタチオンが多量に必要となり、γーGTPもたくさん生成され、血液にまで漏れ出て血液中のγーGTPが増加する。3.免疫力強化1.①グルタチオンはT細胞の活性化とマクロファージのファゴサイトーシス(貪食作用)を維持するのに必要である。②マクロファージやNK細胞などの免疫細胞にグルタチオン量が十分に存在すると、自然免疫力が活発に働き、風邪やインフルエンザにかかりにくくなり、発熱や鼻水、咽頭痛などの症状が軽減する。しかし加齢とともに免疫細胞の中のグルタチオン量は減少して、自然免疫力は低下する。③ウイルスはグルタチオン濃度が高くなると増殖できなくなる。慢性的なウイルス感染症(たとえばAIDS)にはグルタチオン点滴が有効な治療法の一つである。2.①抗原提示細胞であるマクロファージが、抗原をT細胞に提示する際に分泌するIL-12は、ナイーブヘルパーT細胞を、Th1細胞(Thelper1cell)に分化させる。Th1細胞は、IL-2、IFN-γ(IgE抗体の産生を抑制する)、TNF-αなどを産生し、T細胞や単球など貧食細胞の活性を高め、細胞性免疫に関与する。②Th1細胞による免疫応答では、細胞性免疫が働いて、リンパ球やマクロファージなど単核細胞中心の炎症反応が起こる。③マクロファージが抗原をT細胞に提示する際に分泌するPGE2は、ナイーブヘルパーT細胞を、Th2細胞(Thelper2cell)細胞に分化させる。Th2細胞は、IL-3、IL-4(IgE抗体の産生を増加させる。肥満細胞からも産生される。)、IL-5、IL-6、IL-10、IL-13を産生し、液性免疫に関与する。IL-10は、Th1細胞からのIFN-γの産生、IL-12の産生を抑制する。④Th1細胞とTh2細胞は異なるサイトカインを放出することでお互いに抑制し合っているがグルタチオンはTh1細胞とTh2細胞間のサイトカイン反応の正常なバランスを維持する。⑤全体のグルタチオンが減少すると、Th1細胞よりTh2細胞が優位に働き、生体はアレルギー体質に陥りやすくなる。つまり、グルタチオンが欠乏すると、Th1細胞(細胞性免疫の活性化・生体防御)が減少し、Th2細胞(液性免疫の活性化・Ⅰ型アレルギー)が増加するため、感染防御力低下を来たして慢性感染症、慢性炎症が引き起こされ、Ⅰ型のアレルギー(気管支喘息、アレルギー性鼻炎、アトピーなど)が増加する。従って、アトピー性皮膚炎の症状の改善には、グルタチオンを補給することも必要となる。グルタチオン投与は、Th2細胞優位の状態を改善する。⑥逆にTh1細胞が過度に増加すると、細胞性免疫が活性化されて慢性関節リウマチなど自己免疫疾患の発症につながる。
細胞性免疫の活性化 液性免疫の活性化 IgE抗体産生 Th1細胞が過剰に増加すると Th2細胞が過剰に増加すると 自己免疫性疾患 Ⅰ型アレルギー 慢性感染症 慢性炎症 4.抗酸化脂質、抗動脈硬化 ①グルタチオンぺルオキシターゼは過酸化脂質を水、アルコール、 酸化型グルタチオンへと分解する。酸化型グルタチオン 活性酸素やフリーラジカルにより、コレステロールや中性脂肪などの 脂質の過酸化反応が起こると過酸化脂質が生じるが、過酸化脂質 は蓄積すると、細胞膜・細胞機能を障害し、動脈硬化、脳梗塞、心筋梗塞、 糖尿病、がん、老化、認知症、アトピー性皮膚炎、シミ、食中毒などを来たす。 体内での過酸化脂質を減らすには、活性酸素を抑える役割を持つ抗酸化物質 (SOD、GSH、カタラーゼ、ビタミンC、ビタミンEなど)が有効である。 ②動脈硬化は活性酸素による脂質の過酸化と酸化コレステロールを 取り込んだマクロファージが引き起こす炎症が本態である。過酸化脂質 +LH (・OH)ヒドロキシラジカルは脂質LHから電子(e- )とプロトン(H+ ) を奪って脂質ラジカル(L・)が生じ、さらにL・が酸素と結合すると、 脂質ぺルオキシラジカル(LOO・)となる。 このLOO・は近くの脂質(LH)から電子(e- )とプロトン(H+ )を奪って 過酸化脂質(LOOH)になるが、一方電子とプロトンを奪われた脂質は また脂質ラジカルを生じることになり、この反応が連鎖的に繰り返されて、 細胞膜のリン脂質が酸化されつくされると、細胞膜にあるタンパク質が 障害をうけ細胞膜に穴が開き、血管壁の炎症を誘発すると考えられている。 細胞膜のスフィンゴ脂質とコレステロールに富む脂質ラフトにはビタミンE が入り込んでおり、脂質ぺルオキシラジカルLOO-をとらえて過酸化脂質 に変え、自らはビタミンEラジカルとなる。 ビタミンEラジカルは細胞膜に接して存在するビタミンCによってリサイクル されてもとのビタミンEに戻される。 消費されて酸化ビタミンC(デヒドロビタミンC)となったビタミンCは細胞質 でグルタチオンペルオキシダーゼによってもとのビタミンCに戻る。 グルタチオンぺルオキシターゼGPxやカタラーゼはH2O2を安全な水 H2Oに変換することによって、脂質の過酸化の引き金である ヒドロキシラジカル・OHを発生しにくくしている。 ヒドロキシラジカルの産生について ミトコンドリアで生じた過酸化水素H2O2からフェントン反応によって ・OHヒドロキシラジカルが発生するとヒドロキシラジカルは NADPHオキシダーゼ(NOX)という酵素により、体内で積極的 に産生されている。むしろこちらが、メインルートであるとされる。
5.エネルギーレベルの復元、活性化 細胞は24時間、エネルギーであるATPをミトコンドリアで電気エネルギー を利用して生産しているが、常にこぼれ出た電子を酸素が結びついて 副産物である活性酸素を生成している。 ミトコンドリアはエネルギーの製造工場であるが、そこでできた廃棄物 を処理する抗酸化物の1つがグルタチオンである。 このグルタチオンは主に肝臓で作られているが、年齢と共に産生が 低下していきミトコンドリアが発生した活性酸素を処理できなくなる。 そのままではミトコンドリアも細胞も損傷して細胞死に追いやられるが、 グルタチオンを投与することにより、より多くの活力とパワーを提供する ことで、低下したエネルギーレベルを上昇させて、老化や激しい運動 にも耐えられる体力をつける。 6.放射線損傷の保護と修復 ① 直接作用と間接作用 放射線によるDNAの損傷には、放射線が直接DNAを障害する 直接作用と、放射線が細胞内の水と反応をおこして発生した 活性酸素がDNAを障害する間接作用の2つのタイプがある。 ② 直接作用 乾燥状態の物質に放射線を照射すると、放射線エネルギー は標的分子に直接吸収されて、標的分子は励起あるいは 電離し、余分なエネルギーを持つため不安定になる。 この余分なエネルギーを放出する過程で標的分子の 共有結合が切れて2つのラジカルになり、DNAの二本鎖 切断、一本鎖切断をまねく。 ③ 間接作用 放射線のエネルギーは細胞内の水分子に吸収されて、 水分子をイオン化したり、励起したりする。 この水の放射線分解によって・H、・OHの2種類のラジカルと、 H2、H2O2の2種類の分子状生成物及びeaq-を生じる。 これらから次のような活性酸素種が生じる ・O2 (超酸化物陰イオン) ・OH (ヒドロキシラジカル) ・RO (アルコキシルラジカル) ・ROO (ぺルオキシラジカル) H2O2 (過酸化水素) HOCI (次亜塩素酸) 1O2 (一重項酸素) 放射線による間接作用で水分子の電離により産生する 活性酸素のうちで非常に反応性が高く、最も強力な 酸化作用を持つものは、ヒドロキシラジカルである。 生体内における放射線障害の80%はヒドロキシラジカルによる と考えられており、発生するヒドロキシラジカルをどれだけ消し去る ことができるかが放射線損傷によるダメージを少なくする決め手で あると思われる。 そして次のような抗酸化システムによってDNA損傷の保護 と修復が行われる。 SOD CAT (カタラーゼ) GPx (グルタチオンペルオキシダーゼ) チオレドキシン還元酵素 グルコース-6-リン酸 脱水素酵素 現在のところヒドロキシラジカルを選択的に解毒する消去系は 発見されておらず、水素分子がヒドロキシラジカル以外の 活性酸素種(スーパーオキシド、過酸化水素、一酸化窒素) とは反応することなく、ヒドロキシラジカルとのみ反応することを 利用して、水素点滴や水素カプセル(経口)などが試されている。 水素分子は安全で、細胞内の膜でも拡散してすべての オルガネラ(細胞小器官)に到達しうるため効率が良い。 放射線の間接作用によりDNAは二本鎖切断、一本鎖切断 を引き起こすが、ラジカル捕獲剤(例えばR-SHあるいはヒスチジン) により抑制される。 また放射線の間接作用は、塩基の変化を生じさせる。 それには酸素の存在が必要であり、SH基を含む物質(R-SH) により抑制される。 ④ 放射線によるDNA切断の修復 二重らせんのうち一本鎖切断の場合は、DNAの損傷はほとんど修復されるが、 二本鎖切断の場合は、DNAに転座、欠失、重複などの修復ミス(突然変異)が おこることが多い。 活性酸素による塩基の酸化障害に対しては塩基除去修復がうまく行って 元の状態に復帰できる場合と、DNAの複製がうまく行かず塩基置換変異 をおこして、がん化や細胞の機能変化を来たす可能性がある。 7.ミトコンドリア合成の促進 ① グルタチオンは、PGC-1αを活性させてミトコンドリアのDNA 合成を促進して、ミトコンドリアの量と数の増加を来たす。 ② グルタチオンは(PGC-1αを介して)筋肉中のミトコンドリア を増加させることによって、運動機能性の向上を来たす。 ③ 筋肉内でのミトコンドリア新生は、運動時だけでなく、 安静時においても筋肉組織中の代謝を活性化させ体内 の脂肪の取り込みや脂肪の燃焼を効率的に進めて、 脂肪代謝を促進する。 ④ PGC-1αは、血液中のブドウ糖を骨格筋に取り込む 糖輸送体であるGLUT4を増加させて、筋肉組織中の 糖代謝も促進する。 ⑤ 以上のことより、グルタチオンはPGC-1αを活性化し、 ミトコンドリアDNAの産生量を増やすことによって、運動機能 の維持だけでなく、体内の脂肪代謝を促進して、糖尿病などの 生活習慣病を予防する効果を有すると言える。 8.虚血・再灌流障害 虚血に伴う生体組織の虚血障害および虚血後の再灌流 に伴う再灌流障害を虚血・再灌流障害という。 脳梗塞においては、血流の停止によって組織への酸素供給 が絶たれて低酸素状態となり、血流再開によって急激に酸素が 供給されることで酸化ストレスが発生して、血管内皮細胞障害、 微小循環障害をきたし、臓器障害に進展する。 虚血細胞においては、酸素が足りなくなり電流が流れにくく なるため電圧が高くなり、と電子が漏れやすくなる。 そこへ急に血流が回復して酸素が増えると、ミトコンドリア膜 上に電子が急激にあふれて電子はさらに漏れ出しやすくなり、 増加した酸素と結合して、最初の活性酸素スーパーオキシド を発生することになる。 脳虚血再灌流の症状としては、脳内出血、大脳半球局所症状、 けいれん、顔面神経痛、眼球痛、片頭痛様の頭痛、認知機能 障害などを来たす。 両総頸動脈を20分間閉塞後再開通させた一過性脳虚血 モデル(BCCAO)マウスにおいては、慢性経口GSH投与は 脳内GSH濃度を上昇させ、BCCAO虚血によるGSH濃度低下 及びGSSG濃度の増加を改善させ、その結果BOCCAOマウス の記憶・学習機能障害も改善された。 脳梗塞発症後、急性期・亜急性期におけるGSH経口投与は、 脳における酸化ストレスを軽減し、脳機能障害を改善する と考えられる。
こどもの発熱の原因とその対処法
発熱を起こす物質を発熱物質と呼びます。体外から由来するウイルス、細菌、真菌などの微生物自身や菌体成分、あるいはそれらから産生されるサイトカインであるIL-1物質を外因性発熱物質といいます。外因性発熱物質が生体に侵入して主に自然免疫系細胞に働くと、発熱活性を有する物質が放出されます。発熱物質であるIL-1が視床下部の視索前野を活性化し、プロスタグランジンが放出される。発熱物質は血流によって脳に運ばれ、血液脳関門(BBB)が存在しない脳室周囲器官の細胞に作用してプロスタグランジン(PG)E2を産生します。産生されたPGE2は脳組織の中へ拡散し、PGE2受容体を活性化しサイクリックAMP(cAMP)を遊離します。サイクリックAMP(cAMP)は神経伝達物質として体温調節中枢である視床下部にシグナルを伝え、体温のセットポイントを上昇させます。また外因性の微生物由来物質に対する受容体も視床下部内皮細胞に存在し、その活性化はPGE2産生により発熱を起こします。

体温調節中枢である視床下部が刺激されると、交感神経系が活性化され、脂肪組織における代謝性熱産生が上昇し、皮膚内を走る血管の平滑筋が収縮することで、体表面の血流が減少し、体表面からの熱放散が抑制されます。一方、発熱シグナルによる運動神経の活性化は、骨格筋におけるふるえ、熱産生につながります。熱産生促進および体表面からの熱放散抑制の2つの作用によって体の深部温度が上昇します。体深部温を上昇させる生理学的意義としては、体内に侵入した細菌類の増殖至適温度域よりも体温を上げ、それらの増殖を抑える作用、温度上昇による免疫系の活性化を促すことです。解熱鎮痛薬の多くは、プロスタグランジン合成酵素群のなかのシクロオキシゲナーゼと呼ばれる酵素の働きを阻害することで、プロスタグランジンE2の合成を抑制して発熱を抑えます。
発熱物質とは何でしょうか?
脳の視床下部にある体温中枢を刺激して発熱を起こす物質の総称。グラム陰性菌の内毒素などの外来性発熱物質と、免疫機能の一つとして体内で放出される内在性発熱物質がある。パイロジェン。ピロゲン。発熱因子とも言います。
生体内に侵入した外因性発熱物質 exogenous pyrogen は、単球 monocyte、マクロファージ macrophage などの食細胞 phagocytes に取り込まれると、インターロイキン 1interleukin-1 (IL-1)、腫瘍壊死因子 tumor necrosis factor(TNF)、インターフェロン interferon (INF)、コロニー刺激因子 colony-stimulating factor(CSF)、マクロファージ炎症ペプチド macrophage inflammatory peptide-1 (MIP-1)などの内因性発熱物質 endogenous pyrogen を分泌する。
内因性発熱物質は血液-脳関門 blood-brain barrier (BBB)を通過し、脳内でアラキドン酸カスケード arachidonic acid cascadeを経てプロスタグランジン prostagrandin を産生する。これが、体温調節の中枢である視床下部 hypothalamus に働いて体温調節の基準値(セットポイント)を移動させ、熱産生を起こさせる。
アラキドン酸カスケードでプロスタグランジン類を産生する反応はシクロオキシゲナーゼ cyclooxygenase (COX;コックスとよばれる)によって進行する。アスピリン aspirin やインドメタシンなどの非ステロイド系抗炎症薬nonsteroidal antiinflammatory drug (NSAID;エヌセイドとよばれる)はCOXを阻害するので解熱薬として使われます。
非ステロイド系抗炎症薬のなかで酸性を示さないものは、塩基性抗炎症薬 basicnonsteroidal antiinflammatory drug とよばれ、COX阻害作用は弱い。IL-1、TNF、INF、MIP-1などの内因性発熱物質が免疫賦活物質であることから分かるように、発熱は免疫作用を活性化させる。したがって、発熱に対する解熱薬の使用は慎重でなければならない。MIP-1とはマクロファージ炎症性タンパク質(macrophage inflammatory protein、略称: MIP)は、ケモカインと呼ばれる走化性サイトカインのファミリーの1つである。ヒトではMIP-1α、MIP-1βと呼ばれる2つの主要なメンバーが存在し、それぞれCCL3、CCL4と呼ばれる。走化性サイトカインとは、白血球などの細胞を傷害箇所や炎症部位に誘引するサイトカインで、ケモカインと呼ばれます。白血球を呼び寄せるサイトカインです。サイトカインは、細胞から分泌される低分子のタンパク質の総称で、炎症の重要な調節因子として機能します。侵入した病原体に応答して産生され、免疫細胞を刺激・動員・増殖させる働きがあります。ケモカインは、炎症部で大量に産生され、血管内から炎症組織内への白血球の遊走をもたらし、特に白血球の一種である好中球は、微生物の侵入部位(炎症部位)へ素早く遊走して貪食・殺菌を行います。走化性とは白血球などの細胞の遊走過程を「ケモタキシス(走化性)」と呼びます。
発熱が生じるメカニズムとは何でしょうか?外因性発熱物質であるウイルスや細菌毒の刺激が,マクロファージや単球などに作用して,内因性発熱物質であるインターロイキン -1(IL-1)を産生します。IL-1が視床下部の視索前野を活性化し,プロスタグランジンが放出され体温上昇機構が活動します。脳は体温のセットポイントが通常よりも高いと認識して,体温が上昇するように全身に指令を送るのです。解熱薬であるアスピリンは,このプロスタグランジンの合成を阻害するのです。
発熱物質とは,発熱を引き起こす物質である。外因性発熱物質と内因性発熱物質の2種類があります。外因性発熱物質,あるいはToll-like受容体を直接刺激する外因性発熱物質の作用によって外因性発熱物質は体外から侵入するウイルスや細菌、真菌などの微生物やその菌体成分、あるいはそれらから産生される物質です。単球やマクロファージを刺激して内因性発熱物質を産生します。
内因性発熱物質であるインターロイキン-1(IL-1)をマクロファージや単球が産生する。
外因性発熱物質が体内に侵入することで産生される低分子量の蛋白質です。脳内の温度受容細胞の活動変調を介して体温のセットポイントをリセットし、新しい体温レベルを維持するために自律反応が発現します。
外傷や熱傷などにより外因性発熱物質が体内へ侵入すると、次のような一連の反応が起こります。
外因性発熱物質がマクロファージや単球などに作用する。
IL-1が視床下部の視索前野を活性化し、プロスタグランジンが放出される。
脳は体温のセットポイントが通常よりも高いと認識し、発熱を起こす。基本的に微生物やその産生物である。最もよく研究されているのは,グラム陰性細菌のリポ多糖体(一般的には内毒素と呼ばれる)と毒素性ショック症候群を引き起こす黄色ブドウ球菌(Staphylococcus aureus)の毒素である。発熱は,サイトカイン受容体を刺激する内因性発熱物質(インターロイキン1[IL-1],腫瘍壊死因子α[TNF-α],およびIL-6やその他のサイトカインなど)の放出を誘導する生じる。
プロスタグランジンE2の合成が非常に重要な役割を果たすようである。
解熱鎮痛薬(げねつちんつうやく)。発熱にも痛みにも効果が期待できるのは、どちらの場合も体の中で同じ物質が深く関わっているからです。それが「プロスタグランジン(PG)」と呼ばれる生体物質です。
まずは、体温上昇にプロスタグランジンがどう関係しているのか、発熱の仕組みについてみていきましょう。
ウイルスや細菌などの病原体が体内に侵入すると、マクロファージや好中球といった免疫細胞から、こうした異物を排除するために炎症成分サイトカインと呼ばれる物質(「インターロイキン-1(IL-1)」や「インターロイキン-6(IL-6)」など)が放出されます。これらのサイトカインが過剰に作り出され、それらが血流に乗って脳にまでたどり着くと、脳内の血管で「プロスタグランジンE2」(PGE2)というさまざまな生理活性を持つ物質が産生されます。
このプロスタグランジンが視床下部にある「視索前野」という体温調節中枢を刺激した結果、「体温を上げなさい」という指令が出されることに。すると体はそれを受けて、まずは皮膚の血管を収縮させて熱の放散を防ぎます。風邪などの引き始めに寒けがしたり、顔色が悪くなったりするのは、このため。
また、体内にある熱を生み出す力が強い脂肪(褐色脂肪組織)を燃焼させたり、筋肉をガタガタ震わせたりして、体温を上げようとします。こうして発熱が起こるわけです。
発熱の仕組み

痛み成分を増加させるのも「プロスタグランジン」
発熱が感染から体を守る自然な防御反応であるのと同様、痛みもまた体に“危険”を知らせるためになくてはならないものです。痛みが起こることで私たちは体に異常が生じたことに気づき、生命を脅かす危険から体を守ることができます。
その仕組みを三上先生は次のように解説します。
「けがややけどをすると、傷ついた細胞からアラキドン酸という物質が出てきます。そして、それをもとにして作られるのが“プロスタグランジン”という物質です。このプロスタグランジンは炎症を引き起こすと同時に、ブラジキニンという痛みを起こす物質(発痛物質)の作用を増強する働きもあります。こうして生じた痛みの信号が神経を通って脳に届き、『痛い!』と感じるのです」
このように、発熱の仕組みと同様、痛みの場合もプロスタグランジンという物質が関わっているわけです。
プロスタグランジンは多くの女性を悩ませる生理痛(月経痛)にも深く関わっています。
「月経痛は、厚くなった子宮内膜の一部を体外に排出する際に起こりますが、このとき子宮の収縮に関与しているのが、子宮内膜から分泌されるプロスタグランジンです。このプロスタグランジンの量が多いほど収縮が強くなり、月経痛が増します。特に10代では子宮頸管が狭く硬いため、子宮が過剰に収縮して月経痛がひどくなることがあります」と牧田産婦人科院長の牧田和也先生は話します。
外因性発熱物質は、グラム陰性菌の細胞膜成分のLPSである。
外因性発熱物質は、単球やマクロファージを刺激して内因性発熱物質を産生する。
内因性発熱物質は、BBBにCOX-2を誘導して脳内にプロスタグランジンを産生する。
外因性発熱物質は、グラム陰性菌の細胞膜成分のLPSである。
外因性発熱物質は、単球やマクロファージを刺激して内因性発熱物質を産生する。
内因性発熱物質は、BBBにCOX-2を誘導して脳内にプロスタグランジンを産生する。
プロスタグランジンは、体温調節中枢の視床下部に作用してセットポイントを上昇させて発熱を起こさせる。
解熱薬は発熱物質の産生を阻害して発熱を抑える。
発熱は免疫機能を賦活するので解熱薬の使用には注意が必要である。
内毒素の実体は、グラム陰性菌の細胞膜の構成成分であるLPSである。
外毒素の実体は、タンパク質系の毒素である。
内毒素は、発熱作用が強いが、毒作用は弱い。
外毒素は、発熱作用は弱いが、毒作用が強い。
外因性発熱物質と内因性発熱物質
生体内に侵入した外因性発熱物質 exogenous pyrogen は、単球 monocyte、マクロファージ macrophage などの食細胞 phagocytes に取り込まれると、インターロイキン 1interleukin-1 (IL-1)、腫瘍壊死因子 tumor necrosis factor(TNF)、インターフェロン interferon (INF)、コロニー刺激因子 colony-stimulating factor(CSF)、マクロファージ炎症ペプチド macrophage inflammatory peptide-1 (MIP-1)などの内因性発熱物質 endogenous pyrogen を分泌する。
内因性発熱物質は血液-脳関門 blood-brain barrier (BBB)を通過し、脳内でアラキドン酸カスケード arachidonic acid cascadeを経てプロスタグランジン prostagrandin を産生する。これが、体温調節の中枢である視床下部 hypothalamus に働いて体温調節の基準値(セットポイント)を移動させ、熱産生を起こさせる。
アラキドン酸カスケードでプロスタグランジン類を産生する反応はシクロオキシゲナーゼ cyclooxygenase (COX;コックスとよばれる)によって進行する。アスピリン aspirin やインドメタシンなどの非ステロイド系抗炎症薬nonsteroidal antiinflammatory drug (NSAID;エヌセイドとよばれる)はCOXを阻害するので解熱薬として使われる(消化性潰瘍参照)。
非ステロイド系抗炎症薬のなかで酸性を示さないものは、塩基性抗炎症薬 basicnonsteroidal antiinflammatory drug とよばれ、COX阻害作用は弱い。IL-1、TNF、INF、MIP-1などの内因性発熱物質が免疫賦活物質であることから分かるように、発熱は免疫作用を活性化させる。したがって、発熱に対する解熱薬の使用は慎重でなければならない。
代表的な外因性発熱物質は、グラム陰性細菌 gram-negative bacterium 表層のペプチドグリカンとリン脂質層を取り囲む外膜の重要な構成成分で、これをリポ多糖 lipopolysaccharide (LPS)という。LPSは、内毒素 endotoxin ともよばれる。内毒素に対してグラム陽性菌 grampositive bacterium が産生するタンパク質を外毒素 exotoxin という。内毒素と外毒素を比較すると表1のようになる。
プロスタグランジンは、体温調節中枢の視床下部に作用してセットポイントを上昇させて発熱を起こさせる。
エピソームとヘルペスウイルス。
自律的に複製したり染色体に組み込まれたりする可能性のある染色体外遺伝物質を指す言葉として、エピソームという用語が用いられはじめられたが、しかしその後、プラスミドとはいう用語が染色体外の自律的に複製するDNAとして広く使用されるようになった。プラスミドと 1968年にロンドンで開催されたシンポジウムにおいて、一部の参加者はエピソームという用語を放棄することを提案したが、他の参加者は意味を変えてこの用語を使い続けていた。
今日でも、原核生物の文脈において、染色体に組み込むことができるプラスミドのことを「エピソーム」と呼称されることがある。エピソームは複数世代にわたって細胞内で安定して維持される可能性があるが、ある段階ではそれらは独立したプラスミド分子として存在する[36]。一方で真核生物の文脈では、エピソームという用語は、核内で複製される可能性のある、統合されていない染色体外の閉じた環状DNA分子を意味するために使用される[37][38]。ヘルペスウイルス、アデノウイルス、ポリオーマウイルスなどのウイルスがこの最も一般的な例であるが、これらはプラスミドでもある。他の例には、人工遺伝子増幅中または病理学的プロセス(例えば、癌細胞の形質転換)で発生する可能性のある二重微小染色体などの、異常な染色体断片が含まれます。真核生物のエピソームは、DNAが安定して維持され、宿主細胞で複製されるという点で、原核生物のプラスミドと同様に動作する。細胞質ウイルスエピソーム(ポックスウイルス感染症など)も発生する可能性がある。ヘルペスウイルスなどの一部のエピソームは、バクテリオファージ(バクテリオファージウイルス)と同様に、ローリングサークルのメカニズムにより複製を行う。他のものは、双方向複製メカニズム(シータタイプのプラスミド)を介して複製する。いずれの場合も、エピソームは宿主細胞の染色体から物理的に分離されたままである。
エプスタインバーウイルスやカポジ肉腫関連ヘルペスウイルスを含むいくつかの癌ウイルスは、癌細胞内に潜在的な染色体的に異なるエピソームとして維持されており、ウイルスは癌細胞の増殖を促進する腫瘍遺伝子を発現する。癌では、これらのエピソームは、細胞が分裂するときに宿主染色体と一緒に受動的に複製する。これらのウイルスエピソームが溶解複製を開始して複数のウイルス粒子を生成すると、宿主細胞を殺す細胞の自然免疫防御メカニズムが活性化される。

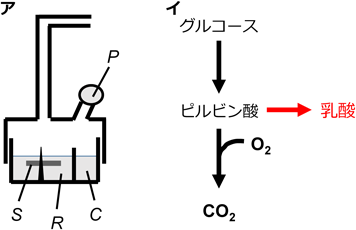{kind=link}
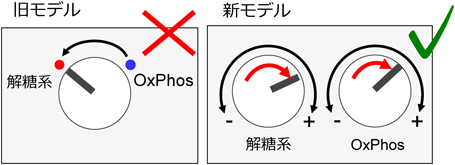{kind=link}
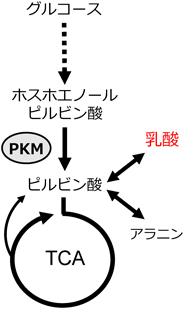{kind=link}
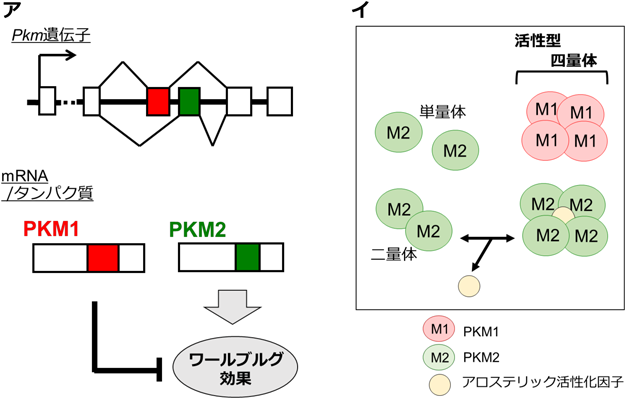{kind=link}
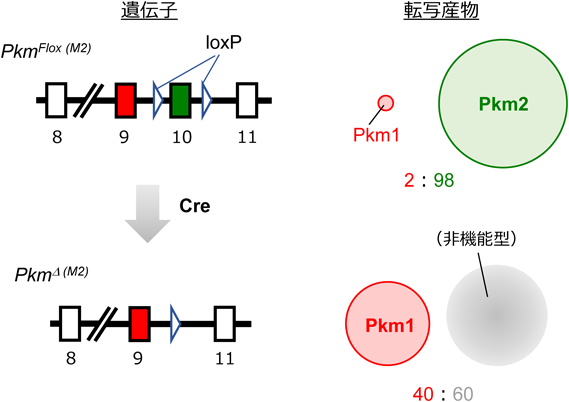{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
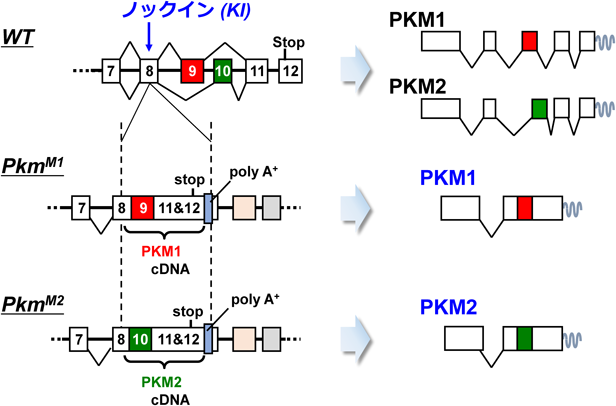{kind=link}
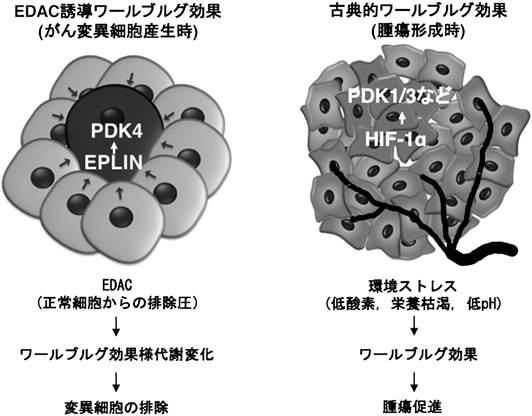{kind=link}
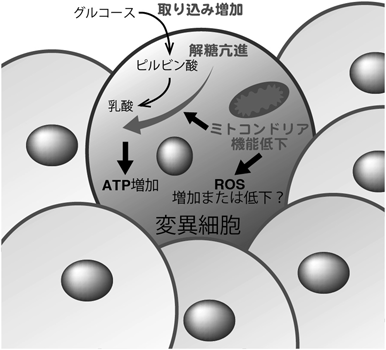{kind=link}
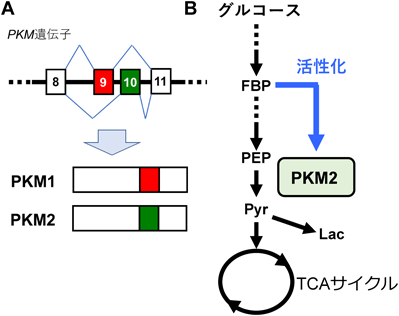{kind=link}
{kind=link}
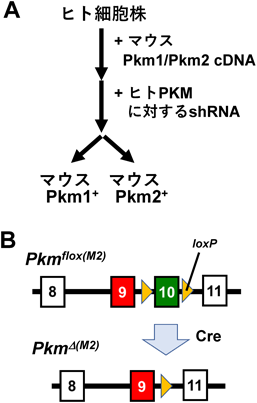{kind=link}
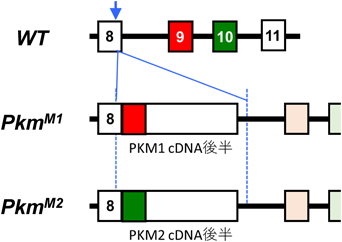{kind=link}
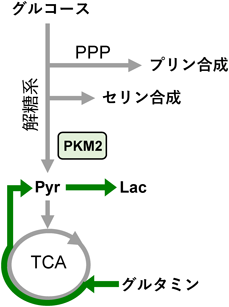{kind=link}
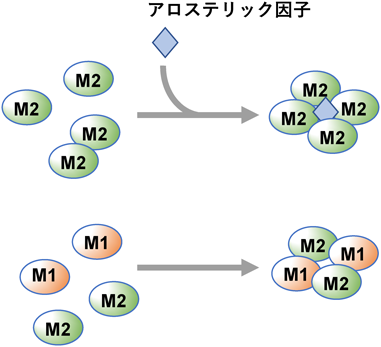{kind=link}
{kind=link}
{kind=link}