何故自己免疫疾患は存在しないのか?自然免疫が自己抗原となるペプチドを敵と認識しないからです。すべての自己免疫疾患は完治できます。何故ならば自己免疫疾患は存在しないので難病といわれる自己免疫疾患は私が自己免疫疾患はすべて治すことができたことが証拠です。治った患者さんの手記や症例報告も完治した証拠として読んでください。
「自己寛容」と 「MHC 拘束」とは何でしょうか?自己免疫疾患は自己寛容のシステムが崩壊すると発症するといわれていますが自己寛容は元々自然免疫が自己免疫疾患をおこさないようにしてくれているので自己抗原寛容は存在しないシステムですから自己寛容のシステムが崩壊する必要もないのです。自然免疫はすべての動物が持っていますが、獲得免疫も併せ持つのは、魚類や両生類、鳥類、哺乳類など脊椎動物の限られたグループだけです。人と同じような免疫システムを持っているラットやマウスなどの脊椎動物が自己免疫疾患を起こすことが有りません。何故ならば魚類や両生類、鳥類、哺乳類など人間と同じく脊椎動物であるので自分たちが持っている自然免疫で自己の成分を自分の敵と見なすことが絶対ないので自己免疫疾患は存在しないのです。もともと存在しない病気が自己免疫疾患ですからその病気の症状を起こす原因はヘルペスウイルスという病原体なのですから自己抗原寛容とは全く無関係な病気なのです。古来から現在に至るまですべての病気は免疫と病原体との戦いであったのです。その間、自己免疫疾患などと言われる病気はなかったのですが俄かに自己免疫疾患という病名が頭の良い金儲けのうまい偉い大学の教授が作り上げたハチャメチャな矛盾だらけの偽の病気として誕生したのです。
人類誕生以来、人類を苦しませてきた病原体はワクチンと抗生物質の発見により征服されてしまいました。病原体でただ一つヘルペスだけがワクチンを作れないウイルスですから人の最後の死に至る癌の原因もヘルペスとなってしまったのです。がんを光療法ですべて治したロイアルレイモンド博士についてはここを読んでください。
自己免疫疾患は存在しないことを完全に理解してもらうために「自己抗原寛容」と「MHC 拘束」の意味を理解してもらうためにも自己寛容とMHC 拘束とは、自己免疫疾患と一体どのように関わり何であるのかの詳しい説明から始めついでに免疫のすべてを復習しましょう。最後に自己寛容と自己免疫疾患とヘルペスとの関わりについて語ることになりますが、結論として自己免疫疾患はないとわかっているのにその誤った理論と長々と付き合うのは面倒ですが正しい免疫の勉強はすべての病気を治すために非常に役に立ちますので自己免疫論者の間違った理論を修正しながら論じていきます。自己免疫論者の間違った理論を完膚なきまで打ちのめして間違った医療で苦しんでいる患者をゼロにしたいのです。
この論文の結論は極めて簡単ですので最初に書いてしまうと「何故、自己のペプチドを乗せたMHC複合体をヘルパーT細胞が敵とみなさないのか?」に対する答えは「自然免疫の敵である非自己だけを認識してくれるTLR」が人体のペプチドを異物である病原体の成分とみなすことができないので自己免疫疾患は存在しないのです。」がこの論文の最終的な疑問に対する非常に簡単な答えとなるのです。TLRについては後述しますが、難解な免疫の根本原理を復習しながらこの答を支える詳しい免疫の論理がこの論文の中核となります。人を非自己である病原体から守ってくれる免疫は学門としても最高に面白い分野ですよ!!!!
自己免疫疾患はない根拠は言い換えると「免疫は元々自分の成分を非自己である病原体から守るために生まれたのであって自己成分を攻撃することは絶対ないのです。それは自然免疫が生まれたのは病原体から守るために生まれ、一方では自分の人体を絶対に攻撃しないシステムを本来持っており、高等免疫である獲得免疫は自然免疫が選別してくれた非自己の病原体だけを効率よく処理できるように進化したのです。敵と味方を判別区別して売れるしてくれる自然免疫があってこそ獲得免疫が自然免疫が選んでくれた非自己である敵だけを殺すことに専念できるのです。自然免疫と獲得免疫との仲立ちをするのが活性化した「樹状細胞」と「大食細胞」と「Bリンパ球」の三つの抗原提示細胞だけなのです。自然免疫がなければ獲得免疫は進化の過程で生まれることはなかったのです。
最も医学的にわかりやすい自己免疫疾患の定義は「自己の主要組織適合遺伝子複合体(MHC)とこのMHCに結合した自己のペプチド抗原の複合体を敵とみなしたT細胞を持っている人です。勿論こんな患者はこの世には誰もいません。
Toll様受容体(Toll Like Receptor、TLR)はヘルペスウイルスや細菌などの人にはない特徴的な構造(分子パターン)を見分けるセンサー(受容体)タンパク質です。Toll様受容体(TLR)は人や動物の細胞表面や細胞内にもある受容体で、ヘルペスウイルスや種々の病原体を感知することができ自然免疫を作動させる機能があり、獲得免疫が働くためにもToll様受容体などを介した自然免疫の作動が人をはじめとする脊椎動物では絶対に必要です。
下図に胸腺の内部構造を示しますが自己免疫論者が言う「正の選択」と「負の選択」は存在しません。胸腺内部は細網組織からなり、多数のリンパ球が密集する皮質と、それよりも明るい髄質が区別されます。細網組織とはリンパ組織に見られ四方へ突起を伸ばす細網細胞と、それが産生した細網線維の網からなる組織のことでつぎの四つの器官に見られる。細網細胞とはリンパ節の支柱を成し,長い数本の突起を伸ばす星形の細胞である.リンパ節ではリンパ球が主体を成す一方,細網細胞が細網線維とともに「網」をつくり,この骨組みの中にリンパ球が容れられる.細網細胞は多彩なケモカインを産生することにより,ニッチの形成に関わっています。
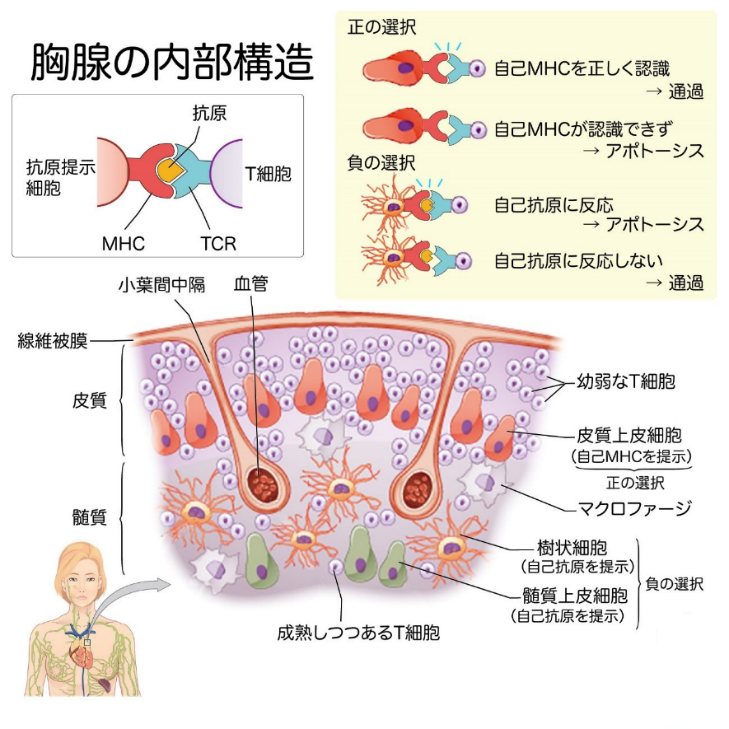
細網組織がみられる4つの器官は①リンパ様組織として総括される組織すなわち、リンパ節、リンパ小節、扁桃、胸腺。②脾臓③骨髄④脂肪組織(これは特殊化した細網組織である。)細網繊維とは細網線維またはレチクリンは、III型コラーゲンで構成される結合組織の線維の種類を記述するために使用されている組織学用語です。細網線維は、網状の繊維が細かい網を形成するために架橋している。この網目は、肝臓、骨髄などの軟組織の支持網やリンパ系の組織や臓器として機能する。レチクリン線維は,網の目を形成して主に骨髄中の支持体の役を果たしています。結合組織中の主な線維状タンパクはコラーゲン線維,弾性線維,細網線維の3つに分けられます。
自己を認識し、自分自身を攻撃しないT細胞を成熟させるために、胸腺では2段階の選抜(正の選択・負の選択)が行われることはありません。あり得ないのですが2段階の選択(正の選択と 負の選択)により自己寛容T細胞を成熟させる仕組みは、まず線維被膜の下で、幼弱な細網組織(プレT細胞)が増殖します。この時に遺伝子組み換えにより様々な形のTCRをもつT細胞がランダムに生まれるのは正常なTCR(T細胞受容体)を形成するためです。
そして、厳しい胸腺学校での2段階の選抜試験が行なわれるのは間違いで行われていませんが間違いを説明しておきます。
まず第一の試験は、皮質にて起こります。皮質上皮細胞が自己MHCを提示します。自己MHCを正しく認識できたT細胞が試験を通過できますのも間違いであり得ません。自己MHCが正しく認識できなかったT細胞達はアポトーシスのスイッチを入れられて、細胞死していきます。この第一の試験を「正の選択」といいますがあり得ません。
第二の試験は髄質にて行なわれます。樹状細胞や髄質上皮細胞が自己抗原を提示します。自己抗原に反応するということは、自分自身を攻撃対象としてしまうことなので、アポトーシスにより排除されていきます。自己抗原に反応しないT細胞が試験を通過できます。この第二の試験を「負の選択」といいます。正の選択も負の選択もされてはいません。
このように、自分自身のMHCがわかることと、自分自身のタンパク質を攻撃しないことの、この2つの条件を満たすT細胞が自己寛容を起こしたT細胞として胸腺学校を卒業できますといわれますがあり得ません。胸腺での選抜は非常に厳しく、2段階の選抜を通過できるT細胞はわずか2〜3%とされていますがウソです。因みに胸腺の上皮細胞と上皮細胞同士をつないでいるのはデスモソームです。デスモソームとはDesmosomeで、接着斑と訳し細胞-細胞間の接着構造の1つです。接着を支える細胞骨格である中間径フィラメントに結合しているデスモグレインやデスモコリンといったデスモソームタンパク質によって形成されています。デスモソームは、細胞膜側面に不規則に配置されたデスモソーム-中間径フィラメント複合体から成っています。
胸腺(thymus)は胸腔に存在し、「獲得免疫」であるT細胞の分化、成熟など免疫系に関与する一次リンパ器官です。胸小葉とよばれる二葉からなっており、胸骨の後ろ、心臓の前に位置し、心臓に乗るように存在する。
胸腺の構造は胸腺は被膜に覆われており、その中に胸小葉がある。しかし胸腺小葉(Lobuli thymi)の解剖学的構造にはまだ定義がありません。この胸腺小葉はさらに外見の違いから皮質と髄質に分けられます。適当な方法で染色すると皮質は濃く染まり、髄質は薄く染まる。
胸腺には以下のような四つの細胞が存在する。①上皮細胞は皮質上皮細胞と髄質上皮細胞の二種類があります。②胸腺細胞はリンパ球です。③大食細胞(マクロファージ)は胸腺内に散在し退化リンパ球や正常なTCRを持っていない出来損ないのT細胞を食べる。④樹状細胞の四つです。

左図の胸腺の顕微写真の説明は白っぽく見えるのが上皮細胞と上皮細胞同士をつないでいるデスモソームです。発生の過程において胸腺の原基は第3咽頭嚢に由来しており、これが下に降りてくる。はじめ胸腺の原基は上皮細胞のみから構成されているが、ここに血流を介してリンパ球の前駆細胞が割り込んで入り上皮細胞を押しのける形で猛烈に増殖する。そのため最初立方形であった原基は伸展・扁平化し網目構造をつくるようになる。
成熟した胸腺において外側の部分である皮質は、上皮細胞が形成する網目の中にリンパ球(胸腺細胞)が詰まっている。このリンパ球は免疫応答をおこせない未熟なものがほとんどである。この未熟なリンパ球を分化・成熟したTリンパ球にするために胸腺があり決して自己の成分や分子に対して免疫寛容を起こすためではないのです。一方、内側の髄質は皮質に比べて上皮間の結合が粗く、リンパ球成分は少ない。しかしこれらは成熟したリンパ球でやがてT細胞として末梢に出ていくものである。髄質にはリンパ球のほかに、マクロファージ(大食細胞)や樹状細胞といった抗原提示の細胞や上皮細胞の変化した胸腺小体(ハッサル小体)が認められる。胸腺小体(ハッサル小体)とは胸腺小体(Hassall’s body略してHC)とは髄質内で特徴的な角化した構造体で胸腺上皮角化小体とも言われます。胸腺小体(HC)を構成している上皮細胞は恒常的に細胞老化を来たし、CXCL5やIL-1ファミリーなどの炎症性分子を高発現している胸腺細網細胞(胸腺上皮細胞)が胸腺髄質において同心円状に配列した領域です。胸腺小体は30~100μm程度の大きさであり、中心には1個あるいは数個の変性度の高い大型細胞が位置する。ハッサル-ウイルヒョウ小体,胸腺上皮角化小体,胸腺小体ともいう.胸腺髄質内にある球状の胸腺上皮細胞の集合したものです。髄質は,皮質と比べて細胞密度が低いが,皮質との本質的な差異は不明である。ただ髄質のところどころにハッサル小体Hassall’s corpuscleとよばれる小細胞集団がみられるのは、変性しかけた上皮性細網細胞が扁平化し同心円状に重なったもので,中心部の細胞は核を失っている。さらにストレスにより分泌された副腎皮質ホルモンや治療で用いられた副腎皮質ホルモンは組織学的には,皮質髄質境界が不鮮明となり,皮質が退縮し,皮質リンパ球が著明に減少し,又,髄質のハッサル小体の占める割合が増大させます。その理由は胸腺は免疫の細胞であるTリンパ球を成熟させる最も重要な一次リンパ器官であるので副腎皮質ホルモンは胸腺における免疫の活動を抑制するからです。胸腺小体(ハッサル小体)は、一言でいうと胸腺髄質内にある変性度の高い核を失っている球状の胸腺上皮細胞の小細胞集団ですが機能は不明です。胸腺小体(ハッサル小体)は細胞の編成集団であって集団であって細胞のないのbodyではないのです。
胸腺髄質にはリンパ球のほかに、マクロファージ(大食細胞)や樹状細胞といった抗原提示の細胞が多いのはこれらの抗原提示細胞(APC)は血流にのって流れてきた病原体の崩壊した成分・分子や自己の細胞のガラクタをとりこみます。非自己である病原体のペプチドや分子に対しては取り込んでPRRであるTLRは異物として認識してAPCとして後に成熟した出来上がったT細胞に異物を提示して免疫の働きを発揮させAPCが正常に働いているかをテストしているのです。正に胸腺は異物を認識できるT細胞を教育してテストする仕事をしており、決して自己免疫疾患を起こさせない教育をする仕事は一切する必要がないのです。
PRRとはpattern-recognition receptorで略してPRRで訳して「パターン認識受容体」です。病原体がもつ特有の構成成分(pathogen-associated molecular pattern略してPAMP)を感知する受容体(pathogen recognition receptor略してPRR)です。PRRは,PAMPを認識後,病原体排除に必要な生体防御機構を発動する.しかし自己の成分やペプチドや分子に対しては異物としてマクロファージ(大食細胞)や樹状細胞は認識してくれないので、つまり、できないので自己成分と戦いは始まりません。何故かというとマクロファージ(大食細胞)や樹状細胞は自己の成分を取り込んでもPRR(パターン認識受容体)であるTLR(トルlike receptor)は自己である自分の成分を危険な異物として認識できる遺伝子の働きが無いので自己免疫疾患は存在しないからです。こんな簡単な真実を世界中の医学者が誰も知らないのは嘆かわしい限りです。存在しないウソの自己免疫疾患を作って治らない病気に仕立て上げお金を儲けて最後は癌にしてしまって人を殺してしまうのです。悲しいですね。自己の快楽だけを追求し続けるのは人間が人間たる所以であるからです。悲しいですね。
胸腺の生理的機能は胸腺では正の選択と負の選択によって適切なリンパ球のみを末梢に送り出しているのは嘘です。正の選択では皮質の上皮に発現する主要組織適合遺伝子複合体(MHC)とこれに結合した自己ペプチド抗原の複合体に対して、適度な親和性があるTCR(T細胞抗原レセプター)を有する胸腺細胞が選択的に増加する。この適度な親和性があるTCR(T細胞抗原レセプター)を有する胸腺細胞が選択的に増加するという表現は一体医学が学問ではないという証拠です。というのは適度な親和性があるという「適度」とは正に「適当な」医学でありウソも適当に含ませているのが職業としての医業なのです。
負の選択では自己MHCと自己抗原との複合体に強い親和性をもつ自己反応性の細胞が髄質内で消去される。強い親和性をもつというのも一体どのように強いのか、かつどのように親和性が測定されその値が高いのか低いのかについては一切説明はないのです。こうした一連の選択は一般に「教育」とよばれるが自己免疫がないような教育も何も必要ではないのです。教育といえば医者たちは病気つくりに専念する暇があれば免疫学の初歩を真剣に復習すべきではないでしょうか?????
T細胞にはキラーT細胞(CTL)とヘルパーT細胞(Th cell)レギュラトリーT細胞(Treg)の三つがあります。キラーT細胞は病原体と感染細胞もろとも破壊する細胞性免疫といわれる働きを専ら行います。ヘルパーT細胞(Th cell)はほかの免疫細胞を手助けするものです。レギュラトリーT細胞(Treg)Tregは、活性化T細胞の働きを抑制します。自己寛容は制御性T細胞(regulatory T Cell略して: Treg) によって制御されますといわれていますが自己免疫疾患はないので、Tregは自己の成分に対して自己免疫寛容を起こすことはありませんし、必要もありません。制御性T細胞(regulatory T Cell略して: Treg)が活躍するのは化学物質や共生可能になった病原体と共存できると判断してくれるのが制御性T細胞(regulatory T Cell略して: Treg)なのです。この時にTreg はインターロイキン10とTGFβというサイトカインを出して戦いをやめさせ病原体と共存する方向に向かうのです。腸管に住んでいる200兆個の共生細菌やヘルペスウイルスとの共存の兆しはあちこちで見られています。PD-1とPD‐L1との関係やCTLA-4とB7との関係がヘルペスとの共存をせざるを得ない兆候となっているのです。PD-1とPDL‐1についてはオプジーボのここを読んでください。
先天的な胸腺不全(ディジョージ症候群など)があると細胞性免疫に欠陥が生じ、感染症にかかりやすくなる。ディジョージ症候群とは何でしょうか?ディジョージ症候群(DiGeorge syndrome)は、遺伝子22q11.2の微細な欠失を基盤とするヘルペスによる先天性の遺伝疾患です。胸腺および副甲状腺の低形成または形成不全による免疫不全・先天性心血管系異常など、さまざまな症状が現れます。また耳介低位や正中線口唇口蓋裂などの特徴的な顔貌を呈します。ディジョージ症候群は胸腺形成不全による遺伝性の原発性免疫不全症の一種です。原発性という言葉は原因不明の病気という意味で用いられます。「原発性」 たいていはヘルペスウイルス感染による先天性又は後天性の免疫遺伝子の突然変異による染色体異常が原因で起こりますが、胎児であるときに生殖性染色体にヘルペスが先天的に感染することは皆無ですが体性遺伝子には胎児のときにヘルペスが感染して胎児のゲノムを突然変異を起こして胸腺の遺伝子を先天的に変えられて生まれた赤ちゃんの先天性の遺伝子病がディジョージ症候群なのです。つまりディジョージ症候群は両親の遺伝子病を受けついだわけではないのです。先天性も後天性の遺伝子病は知らぬ間にヘルペスに感染して発生するのです。
自己免疫疾患が存在しない証拠のひとつがディジョージ症候群は胸腺形成不全にもかかわらず感染症にはかかりやすいのは胸腺は正に分化・成熟したT細胞をディジョージ症候群は作れなくなるからです。同じように胸腺の働きがなくなると自己免疫疾患が増えるはずですが自己免疫疾患にはディジョージ症候群の患者は自己寛容の教育をしていないにもかかわらず絶対にならないのは自己免疫疾患は元々存在しない証拠となるのです。
胸腺の放射線や副腎皮質ホルモンに対する感受性と老化につて・老化胸腺は放射線や副腎皮質ホルモンなどに曝露されると萎縮します。特にT細胞を盛んに産出している10代前後の時期は感受性が高い。胸腺中のリンパ球が最も多いのは思春期(10代前後)でピーク時の胸腺は30 – 40gに達する。胸腺は老化の早い器官で、T細胞を作る役割を担う胸腺が年齢とともに退縮し、T細胞を増やす能力が低下してしまい T細胞を産生する能力は、40歳代で胸腺は脂肪細胞化してしまい新生児の100分の1まで低下します。その後は急速に萎縮し脂肪組織に置き換わってしまうので骨髄で大量に作れた未熟なT細胞の胸腺で自己免疫疾患が起こらない教育がなされないので年取ればとるほど自己免疫疾患は増えるはずなのに減っていくのは何故でしょうか?これも自己免疫疾患がない証拠の一つです。この胸腺の退縮は70歳までにほぼ完了するので70歳以上の人はみんな自己免疫患者であるはずですが新しい患者は増えることはほとんど無いのも完璧な自己免疫疾患はない絶対的な証拠なのです。逆にいえば胸腺は発達が早く、たとえば、出生直後のマウスで胸腺摘出を行うと、マウスは免疫不全に陥るが、成熟マウスで摘出をしても免疫系に影響は少ない。これは、成熟した個体では十分なT細胞のプールができ、末梢でもリンパ球が生理的増殖を行うようになるからであるのはうそです。末梢でもリンパ球が生理的増殖できるからと言われますがリンパ球は非自己の敵である病原体が攻撃しない限り増殖できません。リンパ球が生理的増殖できると聞いたのは初めてです。リンパ球は免疫学的増殖は可能ですが生理的増殖とはいったい何でしょうか?リンパ球が生理的増殖ではなく単一の病原体に刺激されてクローン増殖するのは当たり前ですがリンパ球が生理学的に増殖することも嘘です。リンパ球の増殖は生理学とは一切関りはありません。
血液-胸腺関門とは何でしょうか?胸腺の血管系は胸腺において血管は皮質から入り髄質の方へと向かいますが、皮質側において細動脈・毛細血管は上皮性細網細胞による細胞突起で囲まれているうえ、毛細血管は無窓性の内皮と厚い基底膜をもっており、特にタンパク質は窓のない毛細血管を通りぬけられません。これらの構造はT細胞が分化・成熟する場である皮質にAPCに運ばれてきた抗原以外に余計な抗原が侵入するのを防ぐのに役立っており、血液-胸腺関門 (blood-thymus barrier) と呼ばれる。このように血流による物質の通過を制限する関門として有名なのは血液脳関門(BBB , blood brain barrier)であるが、同様に胸腺もこのような関門によって抗原に会わないように守られているのです。
胸腺の神経については、胸腺の近くには横隔神経や反回神経が通っているので、胸腺の摘出を行う際これらの神経を傷つける恐れがある。横隔神経が麻痺すると横隔膜が上がったままになり息切れしやすくなり、反回神経が麻痺すると嗄声になる。
子供が虐待を受けると子供に大きなストレスがかかり免疫が落ちるので免疫器官である胸腺が高度に萎縮することが知られている。法医学で被虐待児を判別する一つの指標となっている。
過去の西洋医学における胸腺の機能の理解不足と加害の歴史について述べておきましょう。さらに現在も医者たちは新たなる胸腺のウソの働きを編み出して自己免疫疾患という存在しない病気を捏造して、金儲けのために一層悪用して人間に対する加害の歴史に新たなる一頁を加えているのです。1950年代は、一流病院の西洋医学の医師らはX線で子供たちの胸腺を損傷・破壊していた時代でした。幼児期は活発に免疫系を発達させる時期にあたり、骨髄で生まれたばかりの未熟なTリンパ球が分化・成熟させて病原体の抗原を認識させ人間を感染症から守るために胸腺が重要な働きをしていたので、子供の胸腺はもともと肥大しているものなのですが、当時は、胸腺の機能が理解されておらず、幼児期における肥大を何らかの病気の兆候であると誤って判断されていたためである。わずか20数年前までは、”胸腺は子供のときにだけ役目を果たして、大人になると無用のもの”などと、医学部では教えていたのですが、現在でも、胸腺の役目は免疫の司令官である重要な役目をするT細胞というリンパ球を胸腺の中で成熟させることだけが極めて大切な役割であるとわかっているのに何故か胸腺の働きが悪くなると自己寛容の教育ができなくなって自己を敵としてしまう自己寛容ができない異常なT細胞が自己免疫疾患を起こすので治療しないと死ぬよとまで脅かして患者を苦しめまくっているのです。しかも自己免疫疾患は一切存在しないのにもかかわらずです。
自己免疫疾患の症状はこの世で最も恐ろしい永遠に殺し切れないヘルペスであることを隠すためにウソにウソを重ねています。悲しいですね。人体に感染したヘルペスはワクチンができないので従って免疫では絶対に殺し切れないのでヘルペスウイルスは最後は一個の細胞に23500個の内、800個もある癌関連遺伝子を癌化させて癌を作って死にたくない人間を苦しませて殺してしまうのです。人体には40兆個の細胞があるので人体に存在する癌関連遺伝子の総数は一個の細胞には800個あるので40兆×800個もあるので生まれた時の才能の違いも巨大な格差があるにもかかわらず、資本主義は競争社会そのものですからストレスの多い生きにくいウソでまみれている競争社会ではヘルペスが無限大に増えていきますので増えたヘルペスは必ず感染した細胞の核の染色体の遺伝子に隠れて24時間で終わる細胞分裂のたびに何百個、何千個と増えていくので増えたヘルペスが核の遺伝子の癌関連遺伝子の部分的特異的組み換えで遺伝子の突然変異をおこして癌関連遺伝子を次々と癌化させて行くので癌細胞が転移するほどに増えるとヘルペスも猛烈な速度で増えていきます。ヘルペスが増えれば増えるほど800個もある癌関連遺伝子の癌化する確率もうなぎ上りに増えていくので転移癌になると死ぬ以外にないのです。現代のがん医療は活性酸素や放射能や紫外線やたばこ、アルコールがDNAを傷害した結果癌になると極めてあいまいなことを言ってお茶を濁していますが癌を作るのは遺伝子を癌化させるヘルペスなのです。癌についてはロイアルレイモンド博士のここを読んでください。遠隔転移をしない癌はヘルペスが少ないので自己免疫疾患はすべて完治できるように感知できる可能性が出てきました。現代の癌治療は延命が目的ですがヘルペスが大量に増殖してヘルペスが増えすぎて遠隔転移が起こります。ヘルペスが少ない時にヘルペスを増やさない根本治療をやれば遠隔転移は起こらなくなるので癌も治せるのです。今まで何人も治しました。何故またどのように癌は転移するのかについてはここを読んでください。
それではどのようにしてヘルペスウイルスは宿主の癌関連遺伝子を突然変異させて癌化させて癌細胞を生み出すのでしょうか?ヘルペスは感染細胞の核の染色体のDNAに潜伏感染するときには一対のDNAの鎖を切ってヘルペスウイルスのゲノムを組み込んで細胞のヌクレオチドの配列を組み替えてしまうのです。このようなヘルペスウイルスによる宿主のDNAに入り込むときにつかうDNA組み換えを部位特異的組み換えといいます。この遺伝子の部位特異的組み換えによってたまたま2種類の癌関連遺伝子の配列が変わってしまうと一個の癌細胞が生まれてしまうのです。この時細胞のDNA修復機構が部位特異的組み換えを正常なもとの遺伝子配列に修復できなければ確実に一個の癌細胞が生まれてしまうのです。
「獲得免疫」であるT 細胞は、非自己である病原体のペプチドと結合した自己 MHC 分子の複合体としてしか認識できないように拘束されています。何故、病原体のペプチドを単独に敵と認識できる自然免疫が自己だけしかもっていない自己 MHC 分子(HLA分子)に乗せてしか非自己である病原体を免疫システムに知らせざるを得ないのでしょうか?ややこしいですね。
自分の体と同じものを自己、異なるものを非自己といいます。私たちの身体の免疫システムは自己と病原体などの異物などを非自己とを識別して、身体に侵入してくる病原体などの非自己を退治します。 このような非自己だけに応答する機構の中心となるのが細胞性免疫の中心となるT細胞です。そして、ウイルスのような非自己が体内に侵入した場合、それを抗原として免疫細胞が排除してくれるようになるのです。この仕組みを免疫といいます。
免疫は自己と非自己をどのように区別するのでしょうか?自己と肘ことを区別してくれるのは自然免疫のTLRなのです。TLRとは何でしょうか?
Toll様受容体とはToll様受容体 (TLR) は受容体で、マクロファージや樹状細胞に発現し、外来病原体の認識とその応答を担っています。TLRは病原体関連分子パターン (PAMPs) を認識し、自然免疫応答において重要な役割を果たします。ところが自己の成分は外来病原体ではないのでマクロファージや樹状細胞のToll様受容体 (TLR) の受容体では認識されないのです。自分の味方である自分の成分をマクロファージや樹状細胞のToll様受容体 (TLR) が認識しないのを自己免疫論者は気が付いていないのです。TLRは侵入してくる病原体に対する最初の防御システムに関与し、炎症において免疫細胞の生存および増殖を制御しますが自己の成分は全く免疫の敵にはならないのです。
Toll様受容体はどこにあるのでしょうか?
TLRは膜貫通型タンパク質であり、マクロファージや樹状細胞の他にも様々な組織や細胞の種類で発現がみられます。今日までに、11種類のTLRファミリーが同定されており、そのうち、TLR1、TLR2、TLR4、TLR5、TLR6、TLR11は細胞表面に局在し、一方、TLR3、TLR7、TLR8、TLR9はエンドソーム/リソソーム分画に局在しています。
異物を排除する細胞のことを、免疫細胞といいますが人間の成分は異物ではないので免疫細胞には全く関係がないので す。ただし、「異物」を認識するためには、「自分の体と同じもの」と「それ以外のもの」を区別する必要があります。この区別を最初に決めるのは自然免疫であり、自分の体と同じものは自己、異なるものは非自己といいます。この判定を最初に行うのはマクロファージや樹状細胞などの抗原提示細胞(APC)の細胞表面に局在しているタンパク質であるTLRや膜貫通型のTLRなのです。TLRで外来の病原体の関連分子パターン (PAMPs) を認識したマクロファージや樹状細胞などが外来の病原体を貪食して病原体の持つ蛋白を消化し分解して作ったペプチドをAPCが持つMHC 分子(HLA分子)に載せます。このペプチドとAPCが持つMHC 分子(HLA分子)複合体をリンパ球のT細胞に「病原体の分子ですよ」と見せつけるので、「抗原提示細胞(APC)」という名前を付けられたのですが正しくは、「T細胞活性化細胞」というべきです。というのはT細胞には二種類の細胞があり一つはヘルパーT細胞であり、もう一つはキラーT細胞でありますが、このヘルパーT細胞もキラーT細胞も仕事を開始するためにはまず抗原提示細胞(APC)に病原体のペプチドと結合したMHC 分子複合体によって活性化される必要がありますので、マクロファージや樹状細胞はリンパ球活性化細胞と呼ぶべきなのです。
体外から侵入してきた病原体の非自己は抗原とよばれます。人間と同じで意識の中で自分と他人を明晰に分別できるためには自分が自分である「自意識」を確立しなければなりません。同じように肉体にウイルスのような物質である異物が侵入したときに免疫がウイルスは非自己であるかどうかを免疫学的に認識できかつ排除すべき敵であると認識が可能になったのは自己の MHC 分子が誕生したからです。
実は敵のペプチドを自己の MHC 分子に乗せて病原体の分解したあらゆるペプチド成分だけを認識して攻撃できる自然免疫があります。しかし自然免疫だけでは特異的な選ばれたワクチンができないので敵の病原体が侵入するたびに自然免疫だけで同じ病原体とくり返し戦わざるを得なくなると死ぬまで同じ病気を繰り返すことになります。この難関を突破できたのは敵のすべてではなく特定の敵だけを認識し記憶し、二度と同じ病気が繰り返されないように進化し誕生してワクチンを作れるようになった獲得免疫(適応免疫)なのです。
MHC 分子とは何でしょうか? MHCはMajor Histocompatibility Complexの略語であり 主要組織適合性遺伝子複合体とか短く主要組織適合性複合体と訳します。このMHC(HLA)が生まれたのは自己と非自己とを区別するための最も大きな目的は古来から人類を苦しめた病原体を排除するためであったのです。つまり病原体を敵である「非自己」であることを知るために「自分」に病原体が襲い掛かってきたという証拠の「自分の自己証明書」としてのMHC(HLA)分子に敵を結び付けて免疫系のすべての自分のMHC(HLA)分子を持っている仲間の免疫細に安心して攻撃してくださいという「許可証」であったのです。MHC(HLA)分子を主要組織適合性遺伝子複合体と訳していますが実は長くなりますが「主要組織適合性遺伝子蛋白抗原複合体」というべきなのです。何故ならば他人の臓器を移植すると拒絶反応が起こるのは他人のHLA抗原を異物と認識しているから攻撃して排除してしまう拒絶反応が生じてしまうので「主要組織適合性遺伝子蛋白抗原複合体」と明確に言うべきなのです。
HLA抗原は1954年に白血球で発見されたので白血球の血液型と考えられました。HLA抗原はHuman Leukocyte Antigenの略語でヒト白血球抗原と訳します。HLAは遺伝子の第6染色体短腕部に存在する主要組織適合遺伝子複合体(MHC)の産物です。 赤血球を除くほぼ全ての細胞と体液に分布し、自己と非自己の識別に関与する重要な免疫機構として働いていますと言われますが、自己免疫疾患はないので自己と非自己の識別は実は概念としては価値はありますが実際の免疫学では臓器移植以外の分野ではあまり重要ではないのです。
しかし、外部から侵入した細菌やウイルスなどの非自己である病原体を自然免疫の抗原提示細胞(APC)が選別し、病原体由来のペプチドを結合してT細胞に提示することで、免疫の働き手であるキラーT細胞やB細胞などに危険情報を伝えてくれます。元来、免疫にかかわる白血球細胞で発見されたのでヒト白血球抗原(Human Leukocyte Antigen略してHLA)と当初は呼ばれたのですが後になってHLA はMHCと同じものであることが判明したのです。
HLAはヒトのなかで最も遺伝子の多型性(遺伝子の個人差)を示す遺伝子で、白血球の身分証明書の様なもので大きな個人差があり現在、HLA-A、-B、-C および-DRで1万種を超える遺伝子型が知られています。 そのため、個体によって免疫細胞の表面に発現しているHLA分子は非常に多様性に富んでいます。免疫反応に必要な多くのタンパクの遺伝子情報を含む遺伝子領域であり、ほとんどの脊椎動物も持っているのです。つまり脊椎動物も言葉はしゃべれないのですが人の仲間ですから自分が自分だけであると主張しているのです。アッハッハ!
人同士でも臓器移植が簡単に行えないのは80億人の人のMHCがすべて微妙に異なるからです。それはMHCの遺伝子が100%全く同じである人は実は誰もいないからです。従って臓器移植の後必ず拒絶反応が多かれ少なかれ起こるので免疫抑制剤を用いざるを得なくなるのです。
私たちの細胞の表面には、MHCという糖タンパク質が細胞1つあたり10万個も発現しています。人体には40兆個の細胞があるので10万個×40兆個の無限大の数の自分だけしかもっていない自分が自分であるという証明書を一個の細胞の表面に持っているのです。この証明書があるからこそ免疫細胞膜に10万個も発現しているすべて同一のMHCのことをHLA(Human Leukocyte Antigen日本語で訳して、ヒト白血球抗原)と呼ばれるのです。
HLAとMHCの違いは何でしょうか?HLAは発見当初、白血球のみに存在すると考えられていたのですが、白血球抗原としてだけでなく、ヒトの主要組織適合性複合体(MHC; Major Histocompatibility Complex)として知られるようになり、ヒトに関しては、HLAは MHC と同じです。従ってHLAはMHC と同じで体のほとんど全ての細胞表面で発現がみられます。一つの細胞の細胞膜にはMHCタンパク質は10万個もありますが、このMHCタンパク質は免疫細胞によって認識される細胞膜表面のタンパク質なのです。しかもこのMHCには自分の成分であるたんぱく質の分解産物であるペプチドが乗せられていますが適応免疫(獲得免疫)は非自己である異物とは認識されることは無いのです。自分の体と同じものを自己、異なる病原体を非自己といい、幼児期までの成長期に免疫学的な自己と非自己を確立しています。そして、非自己である病原体が体内に侵入した場合、それを抗原として免疫細胞が排除してくれるようになるのです。この仕組みを免疫といいます。
免疫細胞はどのように抗原を認識するのでしょうか?免疫細胞には受容体というタンパク質がついています。受容体が外来抗原と結合した場合、免疫細胞は抗原だと認識することができます。一方、免疫細胞の受容体に結合される抗原の部位も、タンパク質でできており、このタンパク質を、MHCといいます。MHCは、主要組織適合抗原の略で、MHCは、免疫細胞の受容体と結合する部分です。MHCは、ヒトを含めた様々な動物がもつ膜タンパク質の総称です。その中でも、ヒトの白血球がもつMHCを特に、HLAといいます。HLAは、ヒト白血球抗原の略です。つまり人ではMHCと、HLAとは同じものなのです。
それでは免疫細胞の受容体と結合する部分であるMHCを認識するT細胞がもつ受容体を何というのでしょうか?このMHCと結合した病原体のペプチドの複合体を認識しているのが、免疫細胞の表面に存在する受容体です。その受容体はTCRといい、タンパク質でできています。TCRは英語でT-Cell Receptorの略で T細胞受容体と訳します。このTCRはMHCと病原体のペプチドを同時に認識しているのです。
このHLAを規定している遺伝子領域は第6番染色体短腕にあり、タンパク質の構造および機能の違いから、クラスI(HLA-A、-B、-C など)、クラスII(HLA-DR、-DQ、-DP など )、クラスIII の遺伝子領域に分類されています。HLAはヒトのなかで最も多型性(個人差)を示す遺伝子で、現在、HLA-A、-B、-C および-DRで1万種を超える遺伝子型が知られています。そのため、個体によって細胞表面に発現しているHLA分子は非常に多様性に富んでいます。これにより、自己と非自己(外来の細菌やウイルス)を識別し、免疫反応を開始できます。骨髄移植や臓器移植では、白血球のHLA型が一致しない移植片は非自己とみなされるため、HLA型の一致・不一致が治療効果に大きく影響を与えます。一致すれば移植は成功しますが不一致であれば拒絶反応が起きてしまいます。
MHCには二種類ありMHCクラスIとMHCクラスIIです。どちらのMHCにも抗原ペプチドが乗せられるのですが、MHCクラスIに細胞内抗原ペプチドが左の下の図の様に乗せられます。
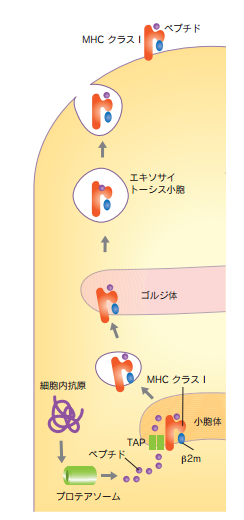
すべての有核細胞は、MHCクラスI分子を持っていて、細胞内のヘルペスウイルスなどの内因性の抗原物質をペプチドに分解し、MHCクラスI分子とともに細胞膜に運ばれキラーT細胞に提示します。またその細胞に感染したウイルスや他の細菌の持っている蛋白質以外に、細胞の持つ構造タンパク質や酵素などの通常の自己の蛋白質をプロテアソームなどの蛋白分解酵素により分解した内因性のペプチドを細胞の表面にすべて提示します。ところがキラーT細胞が非自己の敵と認識できるのは認識できるのはその細胞に感染したウイルスや他の細菌のpeptideと結合したMHCクラスI複合体だけなのです。キラーT細胞も自然免疫である樹状細胞、マクロファージと同じく自己の成分であるペプチドをみなすことができないのです。自己を敵とみなさないので獲得免疫が起こす自己免疫疾患がないという新たなる証拠となるのです。また、免疫細胞である樹状細胞、マクロファージ、B細胞の三つの抗原提示細胞(APC)は、MHCクラスIIも発現していて、外来性の取り込まれた病原体の抗原ペプチドを下の図の様にMHCクラスIIとともにヘルパーT細胞に提示します。MHCクラスIIは外部からふぁごぞーむで取り込んだ非自己の病原体だけが持っている外因性のタンパク質である抗原ペプチドをとのせたMHCクラスII複合体に提示するだけで自己の成分を提示することは無いので自己免疫疾患にはMHCクラスIIは何も関わらないのです。
MHCクラスI分子に結合する細胞内抗原タンパク質(例えばウイルスが感染した細胞内で合成したウイルスタンパク質)は種々のタンパク質分解酵素をもつプロテアソームでペプチドに分解されます。抗原ペプチドは小胞体膜に存在する抗原処理関連トランスポーター(TAP) により小胞体内に運搬されてMHCクラスI分子に結合し、ゴルジ体を経て細胞表面に運ばれます。
小胞体(ER, endoplasmic reticulum)とは何ですか?核の近傍にあって糸状や網目状構造を有する細胞内膜系で、核の外膜との連続性が認められる細胞小器官である。形態は管状、扁平袋状など細胞の種類によって多様である。小胞体は、膜表面にタンパク質合成の場であるリボソームが多数付着した粗面小胞体と、リボソームをもたない滑面小胞体に分けられる。
小胞体とゴルジ体の違いは何ですか?小胞体(ER, endoplasmic reticulum)の役割はタンパク質のゴルジ体への輸送である小胞輸送を行います。小胞輸送とは、膜の分裂や融合により細胞内小器官(オルガネラ)同士あるいは細胞膜とオルガネラの間で、小胞膜を介してタンパク質や脂質などの輸送や、細胞外へ分泌性因子の放出を行う機構です。
真核細胞の細胞内には様々な細胞内小器官(オルガネラ:organelle)が存在しており、これらのオルガネラが機能するためには、それぞれのオルガネラで働く分子が固有のオルガネラへと正しく輸送される必要がある。一方、ゴルジ体(Golgi apparatus/body)とは、ゴルジ装置 、ゴルジ複合体あるいは網状体とも言われ、細胞外へ分泌されるタンパク質の糖鎖修飾をしたり、合成されたタンパク質に様々な糖を付加して糖タンパク質を作り出し、顆粒状にして細胞質に送り出します。付加された糖の種類によって、細胞質にとどまったり、細胞膜に向けて放出されたりします。 すなわち、付加される糖は、そのタンパク質の配達先を記した荷札のような役割を果たします。ゴルジ装置はリボソームを構成するタンパク質の加工を行います。細胞内小器官(organelleオルガネラ)とは何でしょうか?真核細胞の細胞内には様々な細胞内小器官(organelleオルガネラ)が存在しており、これらのオルガネラ(細胞内小器官)が機能するためには、それぞれのオルガネラで働く分子が固有のオルガネラへと正しく輸送される必要があるのです。特に、小胞体(ER, endoplasmic reticulum)、ゴルジ体(Golgi apparatus/body)、エンドソーム(endosome)、リソソーム(lysosome)などのオルガネラ間および細胞表面からのエンドサイトーシスによって細胞内に取り込まれた受容体タンパク質などの輸送は、脂質二重膜からなる小胞膜によって制御されており、小胞輸送(別名、膜トラフィッキング、メンブレントラフィック、細胞内膜輸送など)と総称されます。付加された糖の種類によって、細胞質にとどまったり、細胞膜に向けて放出されたりします。
リボソームとは何でしょうか?真核生物のリボソームは,核小体で転写された rRNA 前駆体に約80種のリボソームタンパク質が結合すると同時に,rRNA 前駆体が加工(プロセシング)を受けるという,非常に複雑 な過程を経て合成される. その合成の場は,核小体から核質へ,さらに細胞質へと移り, 二つのリボソームサブユニットが完成する。リボソームは、数本のRNA分子と50種類ほどのタンパク質からなる巨大なRNA・タンパク複合体です。 全体として大小2つの粒子に分かれ、それぞれ 50Sサブユニット、30Sサブユニットと呼ばれています。 これらのサブユニットの中心部にはRNA分子があり、その表面には多くのタンパク質が結合しています。
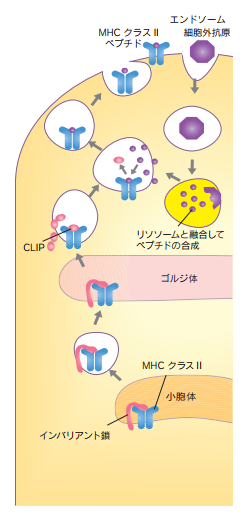
MHCクラスIIに免疫細胞に取り込まれた細胞外抗原ペプチドが左の下の図の様に乗せられます。細胞外の抗原となる細菌、ウイルス粒子、可溶性タンパク質等はエンドサイトーシスやファゴサイトーシスによって細胞内に取り込まれます。エンドソームやファゴソームのような小胞は蛋白分解酵素がつまっているリソソームと融合し、蛋白を分解した結果として細胞外抗原由来のペプチドを生成します。新しくリボソームで合成されたMHCクラスII分子のα鎖とβ鎖はリボソームから小胞体に移動し、蓋の役目をするインバリアント鎖と会合します。抗原ペプチドが膜融合によって入ってくると、インバリアント鎖がはずれ、抗原ペプチドと結合したMHCクラスII分子は小胞輸送で細胞表面に運ばれます。そうすることで、抗原を提示されたB 細胞と T 細胞は、提示されている抗原と結合したMHC ペプチド複合体に注意を集中するようになります。さらに、B 細胞と T 細胞は、自分自身の体を攻撃する可能性のある病原体を体外に排除するために提示された非自己がぴったり合うかどうかを見分ける仕事もしています。一方この世にありえない自己免疫疾患は自然免疫のレベルで除去されているので自己免疫から保護する安全策は多層構造も存在しているといわれていますが自然免疫の大食細胞や樹状細胞が自己の成分は100%安全であることを証明してくれているので自己免疫から保護する安全策の多層構造も必要がないので嘘であり、各層は、上の層で「隙間をすり抜ける」可能性のある自己反応性細胞を捕らえるように設計されているのもウソです。ナチュラル キラー細胞も、自己免疫疾患を引き起こさないことを確認するためにテストされるといわれていますがこれも嘘です。ナチュラル キラー細胞も自然免疫の細胞の一つであり非自己である病原体を殺すことに専念するだけです。
自己免疫疾患を起こさないように「自己寛容」と 「MHC拘束」について膨大な研究が行われてきたものの、まだ多くの未解決の疑問があるのは何故でしょうか?自己免疫疾患がありえない病気なのにない病気が起こるメカニズムがあるわけはないから永遠に解けない未解決の疑問が残ります。いままでの膨大な研究は無駄な労力と時間とお金を浪費しただけです。この世に存在しない自己免疫疾患の病気や治療の研究は即座に中止すべきです。自己免疫疾患の原因はヘルペスですから徹底的にヘルペスのすべての研究に変えればあらゆる病気が完治するのみならず癌も簡単に治ることになるのです。癌についてはライフ博士の「癌療法」ここを読んください。
しかし、この起こりえない自己免疫疾患のトピックを本当に興味深いものにしているのは、それが非常に重要であると自己免疫論者だけが勝手に考えているからです。B 細胞と T 細胞は、この世に存在しない自己免疫疾患のために自分自身の抗原を危険物として認識しないように学習する必要があると言われていますが学習する必要は全くないのです。何故ならばその仕事はそうでなければ、私たちはみな自己免疫疾患で死んでしまうでしょうという愚かな医学者がすべてですが、自己免疫疾患で死ぬ人は誰もいません。しかし正しい免疫学を学ぶことはこの世からすべての病気の原因であるherpesとアレルギーを壊滅するためには価値があります。標準医学の間違いを正しながら免疫学を深めていきましょう。しかも免疫学は医学で一番難解な分野であるに加えてあらゆる病気はお金で治すことができないからです。すべての難病も癌を治すのも正しい免疫でしか治せないので免疫の真実を奥深く知るのは最高に面白いのです。
胸腺とはなんでしょうか?T 細胞はまず、首のすぐ下にある小さな器官である胸腺で自己寛容を学びますと言われていますが間違いです。T 細胞は骨髄で生まれた時に自己を攻撃しない自己寛容を身に着けているのはB細胞や自然免疫の細胞と同じです。すでに自己免疫疾患は存在しない根拠はすでに何回も何回も説明しました。
もう一度繰り返しますが免疫は自然免疫と獲得免疫の二つから成り立っている意味は何でしょうか?自然免疫の一番大事な働きは危険な外来の病原体の非自己の異物と100%安全な自己成分とを正確に見分けるために自然免疫があると言っても過言ではないのです。何故でしょうか?極めて強力な獲得免疫が自分の成分を攻撃させないためなのです。世界中の免疫学者は誰も気が付いていない自然免疫の最も大事な働きは非自己と自己とを厳格に気別するためなのです。そもそも免疫はなぜ生まれたのでしょうか?人体を外来の病原体から守るために進化したのです。進化しすぎた免疫が自己免疫疾患を起こす進化を裏切る戦いを突然起こすことが有りえると思いますか?絶対にあるわけがないのです。自然界を作った神なる造物主は自己免疫を起こさないように自分の成分を攻撃しないように最初に自然免疫に自己は敵ではないので攻撃しては絶対だめですよという教育を施したのです。最初に病原体のすべてを見つけ出す方法と自己成分の見分け方だけを自然免疫に教えたのです。一方賢い獲得免疫には自然免疫だけが認識した病原体だけを攻撃できる術をおしえたのです。教えるためだけに敵と攻撃してはならない自己との見分け方を身に着けたのです。一番簡単な見分け方は病原体である敵だけをPRR (Pattern recognition receptorsパターン認識受容体)であるTLR(Toll-like receptor略して、TLR訳して、トル様受容体)見つけ非自己の敵以外は他は味方ですよと教えたのです。
TLRとは何でしょうか?Toll様受容体(Toll Like Receptor、TLR、トル様受容体)は、細菌やウイルスなどの特徴的な構造(分子パターン)を見分けるセンサー(受容体)で、主にマクロファージや樹状細胞などの自然免疫系の細胞が持っています。TLRは病原体関連分子パターン (PAMPs) を認識し非自己である病原体を取り込んだ大食細胞や樹枝状細胞は病原体が人体に侵入した危険を感知するTLR(Toll-like receptor)といわれるヒトには存在しない細菌やウイルスの構成成分を認識するセンサーを持っています。そのセンサーが感知する細菌やウイルスの構成成分による刺激のことをDanger Signalと呼んでいます。細菌やウイルスが体の中に侵入すると、そのセンサーが感知し防御反応である炎症が始まるのです。
PAMPs(pathogen-associated molecular patterns; 病原体関連分子パターン)とは何でしょうか?PRRs (pattern-recognition receptors:パターン認識受容体)とは何でしょうか?
病原体である細菌やウイルスの非自己の構成成分のことをPAMPs(pathogen-associated molecular patterns; 病原体関連分子パターン)といいます。またPAMPsを認識するセンサーのことをPRRs (pattern-recognition receptors:パターン認識受容体)と総称しています。PRRs (pattern-recognition receptors:パターン認識受容体)とは高等多細胞生物にはない病原体となる微生物特有の分子をPAMPといい、このPAMPを認識する受容体をパターン認識受容体 〔pattern-recognition receptors (PRRs) とよび,PRRs の代表がTLRsなのです。一方,細胞質や核内にも非自己である病原体であるPAMPを認識する分子が存在しRLRs (RIG-Ⅰ,MDA5,LGP2),NLRs (NOD1,NOD2,NALP3など) などが同定されています。これらのPRRsはTLRsとともにPRMs (pattern-recognition molecules) ともよばれる.種々のPRMsは特有のシグナル経路を活性化し,異なった細胞応答に至る。
PRRsによるPAMPの重複認識は免疫応答を多様化し,ゆえに種々の免疫エフェクター細胞の誘導に深く関与する。病原体関連分子パターン(PAMP)は、病原体である微生物内で長く変わることがなく保存されてきた小さな分子モチーフである。糖鎖や複合糖鎖など、さまざまな種類の分子がPAMPとして機能し、大食細胞や樹枝状細胞などの自然免疫細胞に非自己の異物として認識されるのです。
PAMPは、保存されているいくつかの非自己分子で、自然免疫応答を活性化し、感染から宿主を保護する働きと自己免疫疾患を起こさない極めて大切な大切な役割があります。グラム陰性菌の細胞膜上に存在するエンドトキシンである細菌性リポ多糖類(LPS)は、PAMPの原型です。LPSは、自然免疫系の認識受容体であるTLR4によって特異的に認識される。他のPAMPとしては、TLR5によって認識される細菌性フラジェリンや、TLR2によって認識されるグラム陽性菌由来のリポテイコ酸や、TLR2によって認識されるペプチドグリカン、および通常ウイルスに関連する核酸変異体、例えば、TLR3によって認識される二本鎖RNA(dsRNA)、またはTLR9によって認識されるメチル化されていないCpGモチーフが挙げられます。PRRによるPAMPの認識は、インターフェロン(IFN)または他のサイトカインの刺激のような宿主免疫細胞におけるいくつかのシグナル伝達カスケードの活性化をも引き起こすのです。
自然免疫細胞が持つPRR(パターン認識受容体)の代表はTLRなのですがこのPRRは三つの重要な特性を持っています。自然免疫細胞のパターン認識ですから単に一種類の病原体だけを認識するのではなくいろいろな種類の非自己である病原体の共通の一般的な特徴を認識することです。だからこそ自然免疫は特定の病原体だけではなくどんな病原体でも敵として対応できるのです。勿論、自己である自己成分は敵として認識することは絶対にありません。
PRRの二つ目の特性は何でしょうか?PRRが認識する非自己のパターンは病原体にとって非常に重要である特徴であるのでPRRに見つからないように突然変異を起こすことによってそう簡単には変えることができないのです。例えばTLR4が認識するLPS分子(リポポリサッカライド分子)の領域は細菌の外膜を構築するのに必須なのです。したがって細菌がTLR4による認識を逃れるために突然変異をしてしまえば困るのは細菌であるからです。
PRRの二つ目の特性は何でしょう自己成分である分子は認識しないのです。それは認識してしまえば自己免疫疾患で?人類はすべて自己の免疫によって滅びてしまうからです。したがってPRRは自己の成分をDanger Signalと認識することは絶対ないのです。
最近、DAMPs(damage-associated molecular patterns; ダメージ(傷害)関連分子パターン)が体の中にもあったDanger Signalとなる成分が発見されたという愚かな研究者がいます。DAMPsとして機能している分子は平常であるときは細胞内に存在しているのですがヘルペスウイルスなどによって殺された細胞や死にかかっている障害を受けた細胞によって放出された分子なのです。従ってはDAMPsは免疫系に感染によって広範な細胞死や細胞の損傷など,細胞の感染ストレスに伴ってタンパク質から核酸まで放出されますが,細胞の危機を知らせるアラームとしてのみ機能しているだけで、たまたまこれらが非自己である病原体によって自己の成分が崩壊してしまったことを伝えているだけです。
Toll様受容体 (TLR) は敵とだけ結合できる受容体で、自然免疫のAPCと言われるマクロファージや樹状細胞に発現し、外来病原体だけが持っており自己の成分が持っていない分子である病原体関連分子パターン (PAMPs)の認識を行い優れた獲得免疫のTリンパ球に伝える免疫応答を担うのです。TLRは病原体だけが持っている独特な変わることのない分子パターン (PAMPs) を認識し、自然免疫応答のAPCによってTリンパ球に病原体が侵入して来たことを伝える重要な役割を果たします。ところが自己の成分は外来病原体ではないので活性化されたマクロファージや活性化された樹状細胞や活性化されたBリンパ球のToll様受容体 (TLR) の受容体では認識する必要がないのに加えてできないのです。自分の味方である守るべき自分の成分をマクロファージや樹状細胞のToll様受容体 (TLR) が認識しないのでTリンパ球に伝えることができないのです。自己免疫疾患は自然免疫のレベルの疾患ではなく獲得免疫の病気と言い張っているのが自己免疫疾患を唱え続けている愚かな医者たちはこんな簡単な免疫の初歩もご存じないのです。残念です。
このようにTLRは侵入してくる病原体に対する最初の防御システムに関与し、炎症において免疫細胞の生存および増殖を制御しますが自己の成分は全く免疫の敵にはならないのでいかなる自己である成分を危険な分子とみなさないのです。
Toll様受容体は細胞の膜の蛋白質であり膜貫通型タンパク質であります。マクロファージや樹状細胞の他にも様々な組織や細胞の種類でToll様受容体の発現がみられます。今日までに、11種類のTLRファミリーが同定されており、そのうち、TLR1、TLR2、TLR4、TLR5、TLR6、TLR11は細胞表面に局在し、一方、TLR3、TLR7、TLR8、TLR9はエンドソームやリソソーム分画に局在し病原体の核酸も認識できるのです。しかし自己の成分は敵と絶対に認識できないのです。だからこそ自己免疫疾患は出発点からあり得ない病気なのです。
異物を排除する細胞のことを、免疫細胞といいますが人間の成分は異物ではないので免疫細胞には全く関係がないので す。ただし、「異物」を認識するためには、「自分の体と同じもの」と「それ以外のもの」を区別する必要があります。この区別を最初に決めるのは自然免疫であり、自分の体と同じものは自己、異なるものは非自己といいます。この判定を最初6/6に行うのはマクロファージや樹状細胞などの抗原提示細胞(APC)の細胞表面に局在しているタンパク質であるTLRや膜貫通型のTLRなのです。TLRで外来の病原体の関連分子パターン (PAMPs) を認識したマクロファージや樹状細胞などが外来の病原体を貪食して病原体の持つ蛋白を消化し分解して作ったペプチドをAPCが持つMHC 分子(HLA分子)に載せます。このペプチドとAPCが持つMHC 分子(HLA分子)複合体をリンパ球のT細胞に「病原体の分子ですよ」と見せつけるので、APCは「抗原提示細胞(APC)」という名前を付けられたのですが正しくは、「T細胞活性化細胞」というべきです。というのはT細胞には二種類の細胞があり一つはヘルパーT細胞であり、もう一つはキラーT細胞でありますが、このヘルパーT細胞もキラーT細胞も仕事を開始するためにはまず抗原提示細胞(APC)に病原体のペプチドと結合したMHC 分子複合体によって活性化される必要がありますので、活性化されたマクロファージや活性化された樹状細胞や活性化されたBリンパ球は抗原提示細胞(APC)というよりもT細胞活性化細胞と呼ぶべきなのです。
リンパ球の一種であるT細胞前駆体が胸腺に移行し,胸腺で分化・成熟して末梢リンパ組織であるリンパ節や脾臓へと移行する。細菌やウイルスに感染した細胞やがん細胞を除去する免疫系における中心的役割を果たす。このように胸腺は免疫の司令官であるT細胞の産生に必須の中枢リンパ組織であり,髄質および皮質からなる。支持組織である髄質および皮質にはそれぞれ髄質上皮細胞と皮質上皮細胞が存在する。胸腺の二種類の上皮細胞は組織の構築の中心を担うとともに,T細胞を分化・成熟させ成熟したT細胞産生のための主要な支持細胞として機能する.更に中枢性の自己寛容の成立に必須の役割をはたす髄質上皮細胞は,本来,胸腺には存在しない末梢のさまざまな組織に特異的な自己抗原を異所性に発現することにより,これら多くの自己抗原に対し反応しうるT細胞クローンの除去に寄与すると書かれていますがすべて間違っています。自己抗原などは存在しないからです。
はじめに
免疫系は非自己(病原体などの異物)を攻撃しそれを排除するとともに,自己(自分の組織など)を攻撃しない自己寛容の仕組みをもつというのも間違っているのです。正しくは自己のすべての組織にある成分は他人の免疫にとって拒絶すべき抗原となりますが自己の免疫はこの自己のタンパク質・ペプチドは抗原となることは絶対にならないので本来自己寛容の仕組みは必要ではないのです。従って、自分の免疫が自己の蛋白ペプチドに対して寛容することもないのでその寛容が破綻することもないのです。寛容が破綻する自己抗原に対して自分の免疫が破綻したことにより起こるのが自己免疫疾患であるといわれますがこれも嘘です。自己寛容の成立には,リンパ球の発生の過程において起こる中枢性の機構と,成熟したリンパ球が末梢組織において獲得する末梢性の機構とが存在し,この2つの機構は正常な免疫系の確立においてともに重要であるのも嘘です。そもそも自己寛容という現象は存在しないのでリンパ球の発生の過程において起こる中枢性の自己に対する免疫寛容の機構も,成熟したリンパ球が末梢組織において獲得するといわれる末梢性の自己に対する免疫寛容の機構も100%存在しないのです。Tリンパ球の発生の過程で起こることは非自己である病原体だけを処理できる完璧なTリンパ球を産生することと末梢組織においては末梢組織に侵入した外敵を排除することだけなのです。
胸腺組織は病原体を退治する免疫応答の司令塔である病原体を完璧に排除するT細胞の産生に必須の中枢リンパ組織であります。胸腺とは 胸腺は胸骨の裏にある組織で、骨髄で作られた未熟なリンパ球(Tリンパ球)が正常に非自己である病原体だけを殺したり排除するを担っています。 胸腺の機能は幼児期まで活発に働き、思春期で最も大きくなり、その後は年齢とともに萎縮していき、脂肪組織と置き換わることで周囲の脂肪組織と見分けがつきにくくなります。髄質および皮質からなるのは.皮質はおもに自己のMHC分子への反応性の獲得および幅広い抗原に反応しうるT細胞レパトアの形成を担い,髄質は中枢性の自己寛容の成立において中心的な役割をはたす。T細胞レパトアとはT細胞のReceptorが抗原認識の多様性を指すことです。髄質および皮質においては主要な支持細胞として機能する上皮細胞が組織の骨格を形成しているが,髄質上皮細胞は本来は胸腺に発現しないような組織に特異的な自己抗原を発現し,同じ髄質に存在する樹状細胞と協調して未熟なT細胞に自己抗原を提示し,自己反応性のクローンを除去することにより自己寛容の成立に寄与することが知られている。組織に特異的な自己抗原の発現には髄質上皮細胞の発現する核内タンパク質Aireが関与しており,その欠損や異常,および,髄質上皮細胞の分化の異常により,組織に特異的な自己抗原の発現が顕著に低下したヒトやマウスでは臓器に特異的な自己免疫疾患を発症することが知られている。核内タンパク質Aireとは何でしょうか?Aireはauto‐immune regulatorの略であり自己免疫制御遺伝子、自己免疫制御物質と訳します。自己免疫を制御する物質という名前が表すように自己免疫疾患にならないためには必須の遺伝子といわれますがウソです。「自己を攻撃しない」免疫寛容の成立は自然免疫の樹枝状細胞がTLRというPAMPで自己を進化的に敵とみなさい遺伝子を持っているので自然免疫のレベルで自己免疫疾患を排除できているのでAire (auto‐immune regulator)は免疫寛容には全く関りがないのです。Aire (auto‐immune regulator)は自己免疫調節物質, 自己免疫調節遺伝子と訳します。主に胸腺に存在する間質の胸腺髄質上皮細胞に発現し,組織特異的抗原の発現を促すことで自己反応性T細胞を除去することは無いので間違いです。又組織特異的抗原の発現が胸腺髄質上皮細胞に多くなればどうして自己免疫疾患が増えるのでしょうか?疑問です。
これまで,中枢性の自己寛容の成立に必須の髄質上皮細胞を産生する“前駆細胞”は胸腺の発生の過程において同定されていたが,髄質上皮細胞が個体の生涯にわたりどのように維持されるのかについてはよくわかっていなかった。何故ならば人は中年になれば胸腺は脂肪の塊になってしまうので胸腺の自己寛容の成立は不可能になってしまうからです。
更に組織を構成する細胞は,おのおのの組織により,幹細胞,前駆細胞から新たな細胞の産生,成熟した細胞の分裂,のいずれかの機構により維持されるが,髄質上皮細胞のターンオーバーは2~3週間と比較的速いことなどから,幹細胞により維持されているのではないかと想定されてきたが,自己複製能をもつとともに分化した子孫細胞を産生するという幹細胞の定義をみたす細胞は同定されていなかった。
胸腺での免疫寛容を中枢性の自己免疫寛容と言いますがあり得ない現象であると同時に必要性もありませんが論理の間違いを正しながら胸腺の役割をみていきましよう。
脾臓と同様に、胸腺には輸入リンパ管がないため、骨髄から来たCD4もCD8も持っていないdouble negativeのT細胞は血液から胸腺に入ります。しかし、血液中のあらゆるものを歓迎する脾臓とは対照的に、胸腺への細胞の流入はかなり制限されています。何故ならば胸腺は脾臓とは違って病原体を退治する一次リンパ器官ではないからです。骨髄からの未熟な double negative(DN)のT 細胞は、この器官の中央のどこかで波のように胸腺に入ると考えられています。どの本を読んでもこの器官の中央のどこかで波のように胸腺に入るとしか書いていないのは不思議です。リンパ球が血液から二次リンパ器官に出るのを可能にする高内皮細胞(HEV)が一次リンパ器官の胸腺にないため、これがTリンパ球が一次リンパ器官の胸腺にどのように侵入していくのか正確にはわかっていません。わからないことばかりです。
明確にわかっていることは、T 細胞が骨髄から胸腺にCD4も、CD8も発現していないdouble negative(DN)の「裸」で胸腺に入ることです。これらの「裸」のT細胞は胸腺の外側領域である皮質に移動して増殖し始めるのは、「裸」のT 細胞をTCR とかつ正しい補助受容体分子であるCD4か、CD8のどちらかを持たせるためだけなのです。胸腺の役割は正に「裸」の生まれたばかりの「裸」のT 細胞をTCR を持ったかつ正しい補助受容体分子であるCD4か、CD8のどちらかを持った成熟した完璧なT 細胞を育て、非自己の病原体だけを攻撃して殺すための免疫の大司令官にならせるためだけに胸腺の役割があるからです。T 細胞が正常なαβReceptorを持ったTCR(T細胞受容体)が正常かどうかのテストの合格率できるのも、たったの10%未満といわれるのですよ。同時にありえない自己寛容ができるかどうかの非科学的な難解ないくつかにわたるテストにも合格できるわけもないのです。
胸腺は胸骨の裏側で心臓の上に位置し、T細胞が分化・成熟するために必須の臓器であるだけで自己免疫疾患を起こさないための教育は一切必要ありません。骨髄から移入してきた多分化能を有するT細胞の幹細胞は胸腺内の微小環境で成熟し、T細胞へと分化をとげ最終的には末梢へと出ていき、二次リンパ組織や炎症の局所において病原体を殺す機能を発揮します。T リンパ球の分化と選択を担う胸腺微小環境は中年になると脂肪組織になってしまう主に皮質と髄質から構成され,それぞれ皮質上皮細胞と髄質上皮細胞が各微小環境の構造と機能を特徴づけている.
胸腺内におけるT細胞分化の過程で生じる重要な現象は、四つあります。しかしこの中には自己寛容の現象はありません。1)T細胞受容体(TCR)遺伝子の再編成、2)自己MHCを正確に認識するTCRを発現するT細胞の選択である正の選択が可能であるのは、胸腺には皮質上皮細胞、髄質上皮細胞、樹状細胞やマクロファージ等が3次元的に配置され機能的な微小環境が形成されています。APCである樹状細胞やマクロファージが胸腺に存在しているので分化・成熟したT細胞が樹状細胞やマクロファージが取り込んだ非自己の異物に反応した正しいT細胞が選択されて残り、非自己の異物に反応しないT細胞は死にます。APCには樹状細胞の他にマクロファージやBリンパ球の二つがありますがいずれも活性化されないとAPCの仕事はできません。しかしどのようなテストがなされるかは明らかではないのです。合格率は数%しかないでしょう。3)自己抗原を認識するTCRを発現する負の選択であるT細胞の除去は必要ではないのであり得ないので、自己抗原を認識するTCR の負の選択はないのです。4)補助ReceptorであるCD4およびCD8の発現は絶対必要ですが適切なCD4およびCD8であるかはどのようなテストがあるかは不明です。細胞分化の過程はすべて母親と父親の染色体の遺伝子の発現が絡んでいるので出来上がった蛋白質が完全であるかのテストは一筋縄ではいかないので必ず不良品が出るのです。Bリンパ球のB-cell-reseptor(BCR)を産生するときも、母親と父親の染色体の遺伝子の発現が絡んでいるので出来上がったBCR蛋白質が完全であるかのテストは一筋縄ではいかないので必ず不良品が出るのです。この時の正常なBCRの合格率は10%以下だといわれています。
ここで自己のタンパク質成分の分子を遺伝子の進化の中で異物分子とみなさい原理を確立した抗原提示細胞をとは何者なのでしょうか?その原理の結果、自己免疫疾患を起こさせないすごい役割を持っている「抗原提示細胞」(antigen presenting cell)について詳しく説明しましょう。すでに述べたように一番正しい名前はずばり的をついたそのものである「T細胞活性化細胞」(T cell activating cell)です。特に自己免疫疾患ありえないのは「T細胞活性化細胞」がT細胞を活性化できないので今後「抗原提示細胞」ではなく「T細胞活性化細胞」をできるかぎり用いることを宣言します。アッハッハ!世界で初めて「抗原提示細胞」に対して頻繁に用いる医学用語はT細胞活性化細胞」となります。
T細胞活性化細胞はMHC1もMHC2の両方と共刺激とともにT細胞に提供できるのです。共刺激とは、免疫細胞の特にT細胞が、抗原提示細胞の存在下で刺激されて異物である抗原ペプチドを排除するためにT細胞を活性化するためにはT細胞活性化細胞の補助刺激であるB7です。このB7がT細胞のCD28と結合するとT細胞は刺激されることになります。T細胞を完全に活性化するためにT細胞Receptorで敵と結びついて得られる「敵を捕まえた」という刺激と、かつT細胞活性化細胞の補助刺激である共刺激となるB7の2つの刺激が必要なのです。
敵が初めて侵入したときこのB7という共刺激とMHC1もMHC2の両方とも提供できるのはあらゆる組織に敵を見張っている斥候の働きをする活性化樹枝状細胞(DC)と活性化マクロファージと活性化B細胞の三つだけなのです。その三つの中で活性化樹枝状細胞(DC)だけが敵が来たという情報を全身に700個もあるリンパ節にいるヴァージンTリンパ球(ナイーブTリンパ球)に伝えることができるのでT細胞活性化細胞の中で最も重要なのです。ヴァージンTリンパ球(ナイーブTリンパ球)とは敵に出会ったことのないTリンパ球です。この樹枝状細胞(DC)は初めての敵を取り込んでTLRで非自己の病原体が持っている危険な分子を発見してMHCと敵のpeptide複合体を作ってリンパ節にいるヴァージンTリンパ球に危険な敵の侵入を伝えることのできるヴァージンT細胞活性化細胞といってもいいのです。しかも自己のタンパク質を100%安全であるので認識できないので自己免疫疾患がないことを保証してくれる私が大好きな細胞が活性化樹枝状細胞(DC)なのです。ワッハッハ!
樹枝状細胞(DC)とは何でしょうか?単球から分化してできた食細胞で、樹状の触手を持ち病原体を食べ消化します。 その後、病原体の情報をリンパ球に伝えます。ワクチン接種でも重要なのはワクチンは病原体を弱くしただけですから樹状細胞 (DC)に食べられると危険な非自己である蛋白分子であることには変わりはないのでTLRで認識できるのです。 樹状細胞 (DC) は、樹木の枝のような突起を持っているのでその名がつきました。機能はヴァージンTリンパ球(ナイーブTリンパ球)を活性化するための三つの要素である高水準のMHCⅠと MHCⅡの上に病原体のペプチドを乗せてかつ補助ReceptorであるB7の三つを備えています。世界中の免疫学者は故意に見逃しているのですが一番大事な樹枝状細胞(DC)の機能は、自己免疫疾患を起こさないために存在しているのです。もとは血液中の白血球の中の単球という免疫細胞の一部で、組織に出ると樹状細胞 (DC)や大食細胞になり体の中のあらゆる組織に分布しています。
樹枝状細胞(DC)はどの様にまた何が活性化させるのでしょうか?ほかの自然免疫の細胞であるマクロファージが病原体を食い殺そうとするときに腫瘍壊死因子(tumor necrosis factor略してTNF)というサイトカインを放出し、このTNFに樹枝状細胞(DC)は活性化されるのです。さらにウイルスに感染した細胞からは抗ウイルス活性をもつインターフェロンαやインターフェロンβが放出されこの二つのサイトカインによっても樹枝状細胞(DC)は活性化されます。最後には樹枝状細胞(DC)は広い範囲の多種類の微生物の特徴となる分子パターンを認識するために利用されるパターン認識レセプター(Pattern Recognition Receptor、略してPRR)の一つであるTLR(Toll like receptor、訳してトル様受容体,略してTLR)を持っています。このパターン認識receptorによって得られた情報が樹枝状細胞(DC)を活性化するのにもっとも大切な役割を果たします。
成熟したT細胞の特徴を見てみましょう。成熟したT細胞は遺伝子再編成に成功したα鎖とβ鎖(一部の細胞はγ鎖とδ鎖)の二種類の遺伝子産物が二量体を形成し、T細胞受容体(TCR)が細胞表面に発現しています。
95%のT細胞は細胞表面にα鎖とβ鎖のヘテロ二量体(5%のT細胞はγ鎖およびδ鎖のヘテロ二量体を発現している)からなるT細胞受容体(TCR)を発現している。その他に補助ReceptorであるCD4あるいはCD8分子のいずれかを発現し、APC(抗原提示細胞)がからの持っているB7という補助刺激シグナルを受けとるための代表的分子であるCD28などの分子もT細胞は発現しています。
TCRとCD3複合体も必要です。TCRを介して抗原を認識したT細胞は、細胞表面で会合しているCD3複合体を介して信号を細胞内に伝達するのです。
このTCRを介して抗原提示細胞上のMHC(組織適合複合体)分子上に提示されたタンパク質抗原を認識する。抗原を認識したT細胞は活性化し、様々な機能(例えば細胞増殖、サイトカイン産生、細胞傷害活性など)を発揮する。同時にT細胞表面には抗原を認識する時のコレセプター(補助Receptor)あるCD4分子あるいはCD8分子を発現している。最も重要な点は、外界から侵入してくるウイルスや細菌などに由来するペプチド抗原に対して特異的に反応するために、我々の体はそれらすべての病原体に対して反応できる様々な異なる種類のTCRをB細胞の抗体Receptorをと作る時と同じように、前もって準備できるような巧妙なシステムを進化の過程で獲得してきたという点です。
そのために、我々の体はTCRを構成するα鎖やβ鎖の遺伝子を、あらかじめ一つの遺伝子として染色体上に準備せず、それらをいくつかの遺伝子断片(遺伝子セグメント)として染色体上に配置し、それら遺伝子断片(遺伝子セグメント)をまったくランダムに組み合わせることにより1種類のTCR遺伝子を作るように進化してきたといわれる遺伝子再編成を行うのです。例えば1000種類の異なるTCRを作るために、1000種類の異なる遺伝子をあらかじめ染色体上に準備しておく方法に比べ、この遺伝子組み換えの方法(例えばマウスTCRβ鎖の場合にはV断片、D断片、J断片、Cβ断片の4種類の遺伝子の組み合わせになる)は、50×2×12×1個の組み合わせの遺伝子(つまり65個)で、1200種類の異なる遺伝子を理論的には作成できることになり、非常に効率がいい。この遺伝子再編成にはRAG(recombination activating gene)1およびRAG(recombination activating gene)2と呼ばれる二つの遺伝子が必要であるが、この二つの遺伝子は染色体上で非常に近傍に存在しており(種により異なるものの3~15kb程度)、一つの大きなエキソンが全長のタンパク質をコードするというきわめてユニークな特徴を有している。さらに興味深い点は、進化的にRAG1とRAG 2遺伝子は、軟骨魚類以降に突然認められるようになること、およびトランスポザーセの活性を有する事から、RAG1とRAG 2遺伝子は本来我々の持っていた遺伝子ではなく、4~5億年前に外界から侵入してきたトランスポゾン由来の遺伝子であると考えられている。
すでに何回も書いたのですが何故自己免疫疾患はないのかの答えを一言でいうと「自然免疫に選ばれたT細胞が活性化しなければ、どの病原体(抗原)も殺すことができないからです。」言い換えると殺すべき病原体(抗原)を選び同時にT細胞を活性化してくれるのは自然免疫の抗原提示細胞(APC)とかT細胞活性化細胞といわれる大食細胞と樹枝状細胞だけがT細胞を活性化してくれます。自己のタンパク質抗原はAPCに危険な敵とみなされないので絶対に自己を敵にすることは絶対に起こらないのです。
決して自己免疫疾患が起きないように免疫が進化したわけでもないのです。しかし病原体だけを確実に殺すシステムが実は自己免疫疾患を起こらないようにしたともいえるのです。
T細胞受容体遺伝子の再編成について詳しく解説しましょう。
胸腺内おけるT細胞の分化のステップは、細胞表面上のCD4およびCD8分子の発現によりCD4-CD8-(double negative略してDN)期、CD4+CD8+(double positive略してDP)期、CD4-SP(single positive)期、或いはCD8-SP(single positive)期におおまかに区別することができ、左から右へと向かうにつれ成熟していくことになる。
骨髄から移動してきた多能性を有する幹細胞は、胸腺微小環境で、TCRの遺伝子再編成、正の選択と負の選択、CD4およびCD8の発現が誘導され、成熟したT細胞となり、末梢組織へと移動していく。正の選択はあっても負の選択はありません。
TCRを構成するα鎖とβ鎖の胸腺細胞での細胞表面への発現は同時に起こるわけではなく、発現には時期的なずれがある。DN(double negative)期にある胸腺細胞内で、まず二本ある染色体の片側の染色体でβ鎖の遺伝子再編成が生じ、再編成に成功したβ鎖(β鎖選択)はプレTCRα鎖という分子と二量体を形成し細胞表面に発現し、DP(double positive)期へと移行していく。β鎖は片側の染色体でβ鎖遺伝子の再編成が生じると、もう一方の染色体のβ鎖の遺伝子の再編成が抑制され(対立遺伝子排除)、一種類のβ鎖しか一つの細胞には発現していない。次にα鎖の遺伝子再編成が生じ、うまく再編成が生じたものが細胞表面にαβ鎖を発現する。これだけの作業が短時間で正確に行うのは大変なことです。B細胞受容体(BCR)生み出す時もTCRを生み出すのと同じように極めて複雑なので成功率は数%なのです。TCRを作成している間に自己免疫疾患を起こさないように更に煩雑な生産的な作業が可能だと思いますか?無理です。
染色体上に存在する異なった遺伝子断片をランダムに連結することにより、異なる種類の膨大な数のTCRα鎖遺伝子産物が生じる。また、再編成のつなぎめで、塩基の挿入や欠失なども生じることから、多様性は増加することになる。同様の遺伝子再編成は、β鎖でも生じており、α鎖とβ鎖の組み合わせによっても、多様性はさらに増加する。
α鎖遺伝子の再編成はβ鎖と異なり、同時に両側の染色体上でおこり、生産的なα鎖の遺伝子再編成が生じ、その結果として正の選択がおこるまで継続するのですがうまく再編成が生じなかったものは死滅してしまいます。このために異なる二種類のα鎖が一種類の細胞に発現できてしまうことになるのです。T細胞受容体の遺伝子再編成に成功した胸腺細胞は次に、正の選択と負の選択と言う試練をくぐらなくてはならないのですが自己免疫疾患を起こさないために正の選択と負の選択と言う試練は起こらないのです。
この頃に、T 細胞の一部は、TCR の α 鎖と β 鎖をコードする遺伝子セグメントの再配置を開始します。TCR の α 鎖と β 鎖はBリンパ球のBCR(B細胞受容体)の重鎖と軽鎖に当たるとのです。これらのTCR の α 鎖と β 鎖の遺伝子再配置が成功すると、T 細胞は、CD4 補助受容体および CD8 補助受容体だけでなく、T 細胞受容体 (CD3 タンパク質複合体を含む) を低レベルで発現し始め一番大切なTCRを完成していきます。その結果、以前は裸だった T 細胞は、すぐにその表面に CD4、CD8、および TCR 分子で「覆われる」ようになります。これらの T 細胞は CD4 と CD8 の両方を発現するため、二重陽性 (DP) 細胞と呼ばれますが、T 細胞のReceptorはCD4 と CD8の一つだけになるまでまだ完成していないのです。
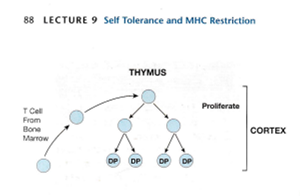
ここまでは間違いはないのですがこの「逆ストリップ」つまり二重陽性 (DP) 細胞の間に、もう 1 つの重要な変化が起こります。ここから証拠もないのに自己免疫疾患を捏造するために牽強付会の間違ったウソの論理展開が始まります。T 細胞がDN(double negative)の裸のとき、つまりCD4 と CD8の二つともないときには、Fas 抗原がT細胞のFasリガンドと結合するとT細胞の細胞死を引き起こす可能性があるのですが Fas 抗原はほとんどまたはまったく発現せず、逆にアポトーシスから保護する細胞タンパク質であるBcl-2を高レベルで発現していたため、アポトーシスによる死に対して抵抗力がありました。しかしFas 抗原がT 細胞のFasリガンドと結合してはじめて、T 細胞のアポトーシスが起こるのですがこの論文では一切Fasリガンドについては触れられていません。何故でしょうか?確かに胸腺皮質の「着飾った」(二重陽性)T 細胞は、その表面に Fas を高レベルで発現していますが、Fasリガンドは全く発現していないからです。一方二重陽性T 細胞はbcl-2 をほとんど生成しません。
Fas (抗原)とは何でしょうか? Fas は、免疫の恒常性を維持するためにプログラム細胞死を媒介する細胞死受容体(アポトーシス受容体)です。 Fas抗原と とそのリガンドである FasL との相互作用により、カスパーゼ 8 が活性化され、その後カスパーゼ 8 がシグナルを送り、下流のエフェクターカスパーゼを介してアポトーシスが開始されます。
bcl-2蛋白とはいったい何でしょうか?胸腺および末梢リンパ組織におけるbcl-2蛋白の発現はどのようになっているのでしょうか?Bcl-2の名称はB-cell/CLL lymphoma 2に由来し、濾胞性リンパ腫における14番染色体と18番染色体間の染色体転座に関与するタンパク質として2番目に記載されたメンバーであることを意味している。CLLはB-cell CLL/lymphoma 2と同義遺伝子名では全部で4つあります。
①BCL 2, ②Bcl-2 , ③Apoptosis regulator Bcl-2, ④B-cell CLL/lymphoma 2、⑤bcl-2、⑥bcl-2CLL lymphoma 2の表記が異なる六つはすべて同じBリンパ球性の慢性リンパ性白血病(CLL)です。慢性リンパ性白血病(CLL)とは、血液のがんの一つで、CLL細胞という異常なBリンパ球が血液や骨髄で過剰に増える病気です。CLL細胞がリンパ節組織で増えている場合には、小リンパ球性リンパ腫(SLL)と呼ばれますが、本質的には同じ病気です。
CLLは英語で(chronic lymphocytic leukemiaで略してCLL)は,訳して慢性リンパ性白血病です。単一な小型円形から軽度の異型を持つBリンパ球の腫瘍で,CD5とCD23の発現がみられ,日本では稀な腫瘍である。慢性リンパ性白血病(CLL)は,表現型的に成熟した悪性Bリンパ球が進行性に蓄積していくことを特徴とする。表現型的本疾患の原発部位としては,末梢血,骨髄,脾臓,およびリンパ節がある。CLLの症状および徴候は,認められない場合もあるが,リンパ節腫脹,脾腫,肝腫大,疲労,発熱,盗汗,意図しない体重減少,早期満腹感などが認められる場合もある。表現型とは、生物の複合的で観察可能な特徴や形質を表す遺伝学の用語であり反対の用語に遺伝子型があります。表現型と遺伝型の違いは何ですか?遺伝子型は、ある生物の個体が持つ遺伝物質のDNAの型です。遺伝子型の変化の違いにより、個体がもつ形態や機能の特徴を「形質」と呼び、遺伝子型に対応する個々の観察できる形質タイプを「表現型」といいます。
CLLの原因は不明であるといわれていますが骨髄でBリンパ球の幹細胞に感染したヘルペスウイルスが,突然変異を起こしたためです。一部の症例では遺伝的要因があるのは妊娠中に感染したヘルペスが原因です。日本と中国ではCLLはまれで,米国の日本人移住者でも発生率の増加がみられないことから,遺伝因子の重要性が示唆されるのは妊娠中のヘルペスウイルスのEBウイルスに感染したためです。CLLは東欧系ユダヤ人で比較的多い。
胸腺リンパ球および末梢リンパ組織(リンパ節および扁桃)のリンパ球のマルチカラーフローサイトメトリーによる解析による次のような結果を得た。フローサイトメトリーとは、乱れの含まない流れ(層流)中で細胞を測定する装置で微細な粒子を流体中に分散させ、その流体を細く流して、個々の粒子を光学的に分析する測定手法のことである。細胞に傷害を与えないで細胞1つずつを一列に並べ、レーザー光を当てて散乱光や蛍光を測定することにより細胞1つずつの情報を取得します。マルチカラーフローサイトメトリーは目的の細胞亜集団にどんな細胞があるかを特定し解析するために複数の蛍光マーカーを使用します。 必要なサンプル数とサンプル量が少ないため、フローサイトメトリー実験の効率が向上します。
胸腺リンパ球および末梢リンパ組織(リンパ節および扁桃)のリンパ球のマルチカラーフローサイトメトリーによる解析による得られた結果は次のようです。
1.最も初期の胸腺リンパ球であるdouble negative cell はbcl-2を表現している
2.次の段階のdouble positive cellは一部にbc1-2を表現していないものがある
3.成熟した胸腺リンパ球であるsingle positive cell はbc1-2を表現している
4.CD1とCD3およびbc1-2による3カラーフローサイトメトリーによると、bcl-2を表現していない細胞はCD1陽性で、CD3を表現していないかごく軽度表現しているリンパ球に限られていた
これらより胸腺ではリンパ球がT細胞リセプターを表現しはじめてpositive selectionを受ける段階の細胞でbcl-2の表現が低下することを示唆している
5.リンパ節および扁桃ではT細胞はすべてbcl-2を表現していたが、B細胞のうち刺激をうけて増殖しているKi67陽性細胞にbcl-2陰性の細胞がみられた。Ki67陰性細胞はすべてbcl-2を表現していた。このことはB細胞においても胚中心でfollicular dendritic cellとの接触によりpositive selectionをうけるB細胞でbcl-2の表現の低下がみられることを示唆している。Ki67陽性細胞とはどういう意味ですか?Ki(ケーアイ)67は細胞増殖の程度を表す指標です。 Ki67陽性の細胞は,増殖の状態にあると考えられています。 したがって,Ki67陽性細胞の割合が高い乳がんは,増殖能が高く,悪性度が高いと考えられるため,より慎重に対処することが望まれます。Ki67とはがん細胞の何ですか?Ki-67は、「がん細胞の増殖力(増殖スピード)」を示す指標です。 細胞が分裂する時に出てくるタンパク質であり、その割合を計測して評価した値です。 Ki-67の値が高い場合はがんの進行が速いことを示し、悪性度が高く再発しやすいと言われています。
以上より、リンパ組織におけるbcl-2の役割は、Tリンパ球のpositive selection と深くかかわっていることが示唆されたといわれますがTリンパ球の「正の選択」には全くかかわりがありません。
bcl2遺伝子は濾胞性リンパ腫のt(14;18)(q32; q21)の18番染色体切断点近傍からクローニングされた。機能的にはアポトーシスに抗する作用があることが判明している。t(14;18)(q32; q21)t(14;18)とは何でしょうか?tはtranslocationの略で転座という意味です。転座により、14番染色体q32のIgH遺伝子と18番染色体q21のBCL2遺伝子がIgH/BCL2キメラ遺伝子を形成するという意味です。キメラ遺伝子とは2つの異なる遺伝子、もしくはその一部が融合した遺伝子をキメラ遺伝子と呼びます。 白血病でしばしば認められる染色体転座(ある染色体の一部又は全部が別の染色体に結合すること)によりキメラ遺伝子が形成された後、正常な細胞には認められない異常なタンパク質が生成され、白血病発症に関わるのです。
転座は、濾胞性リンパ腫(FL)やびまん性大細胞型Bリンパ腫(DLBCL)に特徴的に見られる染色体異常である。転座とは染色体異常の一種。 染色体の一部が正常の位置から別の位置や他の染色体へ移動する現象。 また、二つの相同染色体間で互いに一部の座位を交換する場合もあり、これは相互転座という。転座により、14番染色体q32のIgHと18番染色体q21のBCL2がIgH/BCL2キメラ遺伝子を形成する。本検査は、濾胞性リンパ腫(FL)や瀰漫性大細胞型Bリンパ腫(DLBCL)の診断時に有用なのです。
BCL2蛋白は濾胞性リンパ腫の大部分の症例で高発現しており、一方反応性の胚中心細胞では発現を欠いているので(mRNAは検出できるので胚中心では特殊な制御機構があると推測される)両者の鑑別をするうえで重要な所見である。胚中心細胞とは胚中心はgerminal centerと言い略してGCで、Bリンパ球が成熟するために二次リンパ組織(リンパ節)につくられる微小構造が胚中心で、その胚中心にいる細胞がBリンパ球です。抗体を産生するB細胞の活発な増殖,選択,成熟と消失がみられる部位が胚中心です。 その構造学的特徴から,暗領域,明領域に分けられ,B細胞の成熟過程は主に濾胞樹状細胞(FDC)が存在する明領域で行われる。
B細胞リンパ腫では濾胞性リンパ腫に限らずマントル細胞リンパ腫、MALTリンパ腫などの低悪性度リンパ腫のほとんどで検出されるし、瀰漫性大細胞型B細胞性リンパ腫でも多くの症例で認められる。また、T細胞性リンパ腫や骨髄球系の白血病でも広く発現が見られる。濾胞性リンパ腫では大体8-9割が陽性になるが、陰性例も見られる。興味深いことに、細胞が大型化した、特にGrade3bの濾胞性リンパ腫では陰性例が多い。また、バーキットリンパ腫では大半が陰性であり、瀰漫性大細胞型B細胞性リンパ腫との鑑別点のひとつである。
アポトーシス抑制分子であるBCL-2は様々な細胞に発現し、DNA損傷を始めとする種々のアポトーシス誘導シグナルから細胞を守っております。癌細胞においてBCL-2はしばしば強発現しており、癌細胞の生存に重要な役割を果たしているので癌細胞がアポトーシスしないのです。
その結果、アポトーシスによる死を引き起こす可能性のある信号に対して非常に敏感になります。この非常に脆弱な状態で、T 細胞は MHC 制限と自己寛容についてテストされます。どちらかのテストに合格しないと、恐ろしい死を遂げます。このようなテストはないので嘘です。
この自己寛容というあり得ないテスト以上に胸腺の役割にはもっと大切な成熟したT 細胞になるための他に二つの大きな極めて大切な関門(テスト)を突破しなければなりません。一つ目はαβT細胞受容体(TCR)かγδT細胞受容体(TCR)のどちらかを選んで生み出す必要があります。まずT細胞のRAG1蛋白とRAG2蛋白が染色体の二重鎖を一本鎖に分けて一本鎖の遺伝子をスプライス(再結合)開始します。この時遺伝子をαβT細胞受容体の遺伝子にするかγδT細胞受容体にするかの競争が生まれます。完璧なαβT細胞受容体になるか完璧なγδT細胞受容体になる遺伝子選びの競争の勝率は10%にも満たないのです。このどの遺伝子が選ばれるかの正確なメカニズムは全部解明されているわけではないのですが、95%以上のT細胞がαβT細胞受容体を持っています。残りのγδT細胞受容体が5%です。
この一つ目の関門はT細胞受容体(TCR:T-cell receptor)遺伝子の再構成と言いますが、難解なのでもう一度これを別の切り口で説明しておきます。T細胞は胸腺における発生の過程でT細胞受容体(TCR:T-cell receptor)遺伝子の再構成を起こすことによって1億種類にもおよぶ抗原特異性を獲得します。つまりTCRバリエーションの獲得は,いわゆるゲノム編集によって引き起こされる不可逆的な現象であり,生体内でもリンパ球に限って観察されるのです。具体的にはTCRα鎖とTCRβ鎖をコードするTCRA遺伝子ならびにTCRB遺伝子を再構成するために,それぞれのV遺伝子とD遺伝子とJ遺伝子セグメントがありそのセグメントに様々な多様な遺伝子候補群が集まっています。例えばTCRAではVαとJα遺伝子から,TCRBではVβとDβとJβ遺伝子からランダムにセグメント遺伝子が選ばれ,1つひとつの胸腺内T細胞につき1〜2種類のTCRα鎖と1種類のTCRβ鎖が産生されるようになります。このTCRα鎖とTCRβ鎖の組合せとセグメント遺伝子接合部に入った変異によって抗原特異性が決定されるので一億種類の抗原を認識可能となるのです。Bリンパ球もB細胞受容体(BCR)を1兆種類にもおよぶ抗原特異性を獲得するためにも同じことをやってBCRの多様性を生み出しているのです。
二つ目は補助受容体であるCD4だけを持つヘルパーT 細胞になるかCD8補助受容体だけを持つとキラーT細胞になるかの競争が待ってます。しかもどちらのTも細胞の表面にMHC分子に乗せたペプチドから成り立った複合体を認識できるT細胞を間違いなく作る仕事があります。ヘルパーT 細胞にはⅠ型のMHC分子をキラーT細胞にはⅡ型のMHC分子を載せるようにします。従って胸腺に最初に骨髄から到着したT細胞受容体にかかわる分子を一切持たない完璧に未熟なT細胞は成熟したT細胞になるために胸腺に来たのであって自己免疫疾患を起こさないために胸腺という中枢の免疫器官に来たのではなく外来の病原体を駆除するための教育を受けるためだったのです。ヘルパーT細胞は、胸腺で前駆細胞から発生・分化を遂げますが、この過程で抗原を感知するT細胞受容体(TCR)がランダムな遺伝子組み換えによって数千万種類も形成され、外からのさまざまな病原体の排除に備えます。抗原を感知するT細胞受容体(TCR)がランダムな遺伝子組み換えによって数千万種類も形成され、外からのさまざまな病原体の排除に備えるのですが、抗原を正確に感知するT細胞受容体(TCR)がランダムな父親からの遺伝子と母親からのに遺伝子を遺伝子組み換えによって数千万種類も形成されるのも大事業なのです。ここでうまく、抗原を正確に感知するT細胞受容体(TCR)を作れなかったヘルパーT細胞はアポトーシスで死んでいきます。生き残った完璧なαβT細胞受容体(TCR)を持ったT細胞は病原体の非自己のペプタイドと自己のMHC分子の両方ともを同時に認識できるようになって胸腺から出ていくのです。非自己のペプタイドだけとか、自己のMHC分子だけを認識するのでは二つを認識して初めて非自己を敵として認めるのですよ。
自己と非自己を見分けるために胸腺で教育を受けて「自己抗原に対して免疫寛容」を身に着けかつ自己のペプチドをのせたMHC複合体が自己免疫疾患を起こさないようにするために胸腺があるのではないのです。わけのわからないテストに合格しないと、恐ろしい死を遂げると脅かすのは医薬業界の繁栄のためであって患者は苦しむばかりです。悲しいですね。Fas 抗原とかBcl-2とかが意味を持つのは、まともなTCR(T cellレセプター)を完備した成熟したT細胞がレセプターで得た敵に情報を核の遺伝子まで送ることができるシグナル伝達がかんりょうするまでの活性化された後のいわば高等免疫である「獲得免疫」にまで成長してからの話です。
Fas 抗原とBcl-2が含まれている文章の中に「可能性がある」と言われる話はほとんどが真実であると確信の持てない話であり従ってすべて可能性がないのであり間違った論理なのです。何故研究論文には「可能性がある」という但し書きが多い理由はその可能性がある考えには確たる証拠がないからなのです。私にようにすべての自己免疫疾患を完治させている医者にとっては「自己免疫疾患がある」可能性が間違っていることを100%証明することほど簡単なことはないのです。それをさらに検証していきましょう。しかも自己免疫疾患がないのに何故自己免疫疾患と言われる病気に症状が出るのでしょうか?私は何万人の自己免疫疾患をすべて完治させたのですがその症状の原因はherpesですとまで明確に答えを出しているのです。だからこそ自己免疫疾患と言われる患者に初めて診察する時に患者に伝える最初の言葉は「必ず治してあげる」が口癖になっており実際に抗ヘルペス剤と漢方煎剤と共に投与して完治させてしまうのです。勿論病気を完治させるのは患者さん自身の面絵杭であることは言うまでもないことですが。症例報告や、完治した患者の喜びの声はここを読んでください。
FasやBcl-2の医学用語が出たついでにFasやBcl-2について詳しく説明しましょう。
まずFas 抗原の本当の働きは何でしょうか?がん細胞やウイルス感染細胞の除去にもFasは関わっており、がんの抑制にも機能しているのは本当です。Fas 抗原に対するFasリガンドはアポトーシスを誘導するサイトカインのdeath因子(デス因子、死因子)であります。近年、デス因子(死因子)が次々と発見されているが、FasリガンドはTNF(腫瘍壊死因子)とともにデス因子のプロトタイプ(原型)として最も盛んに研究されている。様々なアポトーシス誘導系の中で、細胞外のシグナルからカスペース(カスパーゼ)の活性化までの経路が明らかになっているのは、デス因子によるアポトーシス誘導系のみである。また、Fasリガンドは細胞傷害性T細胞(CTL)の主要なエフェクター分子の一つとして働いている他、免疫系の制御にも重要な役割を果たしていることが明らかになっている。免疫細胞のエフェクター分子とは又細胞障害性T細胞によるウィルス感染細胞や移植片等の標的細胞にapoptosisを誘導することも仕事です。エフェクター機能とは免疫学で何ですか?抗体のFc領域が担う機能で,抗体クラスに大きく依存している。補体を活性化する機能はIgMとIgGクラスの抗体に限られ,抗体の可変領域が結合した細胞を溶解させる機能を特にCDC(補体依存性細胞障害)と呼ぶ.エフェクター細胞とは何ですか? まず、抗原刺激により、ナイーブB細胞、ナイーブキラーT細胞、ナイーブヘルパーT細胞が抗原特異的に活性化され、増殖します。この 増殖した時期の仕事ができるようになった細胞はそれぞれの役割を果たすべく一生懸命に働きだします。 この時期の細胞を各細胞の「エフェクター細胞」とよびます。
T細胞が活性化するとその表面にFasリガンドが発現する。T細胞はクローン性増殖の間はFas誘導性アポトーシスに耐性であるが、活性化されるとその期間の長さに従ってFas感受性が高くなり、最終的に活性化誘導細胞死(activation-induced cell death略して, AICD)に至る。
細胞傷害性T細胞(TCL)の活性には二種類あり一つはFas誘導性アポトーシスと二つ目のパーフォリン経路は外来性の抗原を提示している細胞を細胞傷害性T細胞(TCL)がパーフォリンが誘導して細胞死をもたらす二大経路である。
免疫特権とは角膜や精巣等の免疫から逃れられる特権領域に存在する細胞はFasリガンドを発現し、浸潤リンパ球のアポトーシスを誘導する。これは、身体が免疫特権の確立と維持に用いている多くのメカニズムの一つである。免疫特権とは何か?全身の免疫系から隔絶されている,臓器内で自己制御されているなどの理由により免疫応答や炎症反応などが起こりにくい性質のこと.免疫特権があると臓器移植の際に拒絶反応が起こりにくいことから移植成功率が高いとされる.脳,眼(角膜),毛髪,精巣,母親の子宮内の胎児などに免疫特権があると考えられている。免疫特権とは何でしょうか?免疫特権(immune privilege)とは全身の免疫系から隔絶されていると考えられ,かつ臓器内で自己制御されているなどの理由により免疫応答や炎症反応などが起こりにくい性質のことで、免疫特権があると臓器移植の際に拒絶反応が起こりにくいことから移植成功率が高いとされる.脳,眼(角膜),毛髪,精巣,母親の子宮内の胎児などに免疫特権があると考えられている.何故免疫特権が起こるのか?免疫系は、病原微生物などの異物を速やかに排除し、生体を守るために進化した必須の防御システムですが、脳,眼(角膜),毛髪,精巣,母親の子宮内の胎児などの需要な臓器における補体を含む自然免疫も獲得免疫も免疫応答は、正常組織も攻撃するリスクをはらんでいるとも言えます。 このため、生体には免疫監視機構が発動しにくい組織や空間が存在しており、これらを「免疫特権部位」と呼びます。免疫監視機構とは何ですか?免疫とは、細菌やウイルスなどの外敵が体に侵入してきた時に敢然と立ち向かい退治し、身体を守ってくれる働きや仕組みのことをいいます。 そういう仕組み全体を免疫監視機構、生体防御システムといい、そしてその重要な役割を担っているのが、血液中の白血球です。
次にBcl-2とは何でBcl-2の役割は何でしょうか? bcl-2は、アポトーシス抑制活性を持つがん遺伝子である。Bcl-2ファミリータンパク質の主要機能は、ミトコンドリアにおけるアポトーシスの制御である。 紫外線やγ線照射、また活性酸素による酸化ストレスなど種々の刺激は、BAX及びBAKの活性化を通じてアポトーシスを引き起こす。γ線照射とはガンマ線は非常に透過力の強い光子であり、コンクリート壁や厚い銅板でなければ遮断することができません。 この高い透過力を利用して、身体の奥にできたがんの治療に応用。 そのほかには、医療廃棄物や食品の滅菌などにも利用されています。放射線であるガンマ線照射による滅菌は、温度上昇もなく、安全で確実に菌を死滅させることができる滅菌方法です。 ガンマ線は透過力に優れているため、梱包形態の制限も、線量のバラツキも少なく、金属や水入り製品粉末、フィルム原反など高密度製品も滅菌可能です。原反とは「げんたん」と読原反やロール紙とは、まだ加工(裁断)されていないロールの状態のことを指します。 紙に限らず、ロール状のフィルムや金属箔のことも同様の呼び方をします。BCL-2はアポトーシス促進性タンパク質(BAX/BAK、BIMなど)と相互作用することにより、アポトーシス抑制性に機能している。BAX、BAK、 BIMとは何でしょうか? Bakはミトコンドリアに局在しアポトーシスを促進するタンパク質です。BAK、BIMもアポトーシス促進性タンパク質です。 Bcl-2ファミリーのメンバーです。
Bcl-2(B-cell lymphoma 2)はヒトではBCL2遺伝子にコードされるタンパク質で、細胞死(アポトーシス)の阻害または誘導のいずれかを行う。Bcl-2 は、B-cell lymphoma の略で、1985年にヒト濾胞性リンパ腫から単離された癌遺伝子としてBcl-2遺伝子が発見されたことからこのように名付けられました。Bcl-2ファミリーの最初に発見されたメンバーである。このファミリーの中には、Bcl-2、Bcl-XL、Mcl-1 などアポトーシスを抑える働きをするタンパク質がある一方、Bax、Bak、Bad、Bim などプログラム細胞死(Programmed cell death)へと続くカスケードのトリガーとして機能するタンパク質もありますからややこしいですね。いずれも細胞の生死に対して極めて大きい影響を及ぼしており、「アポトーシス・チェックポイント・ファミリー(Apoptosis checkpoint family)」とも呼ばれます。Apoptosis checkpoint familyとは何でしょうか?アポトーシスを行うべきかどうかを注意して確認すべき点や箇所に関わるたんぱく質の仲間。BCL-2はアポトーシス促進性タンパク質(BAX/BAK、BIMなど)と相互作用することにより、アポトーシス抑制性にも機能しているわけですからアポトーシスの阻害または誘導のいずれも行うことができるのがBCL-2なのです。
アポトーシス促進性タンパク質(BAX/BAK、BIMなど)とは何でしょうか? Bcl-2(B-cell lymphoma-2)ファミリーのタンパク質は癌の領域で、30 年以上に渡って研究が続けられてきました。それはこのファミリーの中には、Bcl-2、Bcl-XL、Mcl-1 などアポトーシスを抑える働きをするタンパク質がある一方、Bax、Bak、Bad、Bim などプログラム細胞死(Programmed cell death)へと続くカスケードのトリガーとして機能するタンパク質もあり、全く相反する生と死に関わっているからです。いずれも細胞の生死に対して極めて大きい影響を及ぼしており、「アポトーシス・チェックポイント・ファミリー(Apoptosis checkpoint family)」とも呼ばれているのです。
アポトーシス・チェックポイント・ファミリーとは何でしょうか?アポトーシスには外因性と内因性の2つの経路があり、内因性アポトーシスはBCL-2ファミリータンパク質により制御されている。BCL-2ファミリータンパク質は、以下の3つのタイプに分けられる。
①アポトーシス抑制タンパク質:BCL-2、BCL-XL、BCL-w、MCL-1など
②エフェクタータンパク質(アポトーシス促進性タンパク質):BAX、BAKなど
③BH3-onlyタンパク(アポトーシス促進性タンパク質):BIM、BCL-XS、NOXAなど
内因性アポトーシスでは、BAX/BAKがミトコンドリア外膜上でオリゴマー化して細孔を形成し、ミトコンドリア外膜透過性が亢進する。これにより、チトクロムcがミトコンドリアから細胞質へと漏出し、カスパーゼ-3/7などが活性化され、アポトーシスに至る。
BIMなどのBH3-onlyタンパクは、DNA損傷、細胞傷害などが起こると活性化され、BAX/BAKなどのエフェクタータンパク質を活性化することで、上記のアポトーシス経路が進行する。
一方、BCL-2などのアポトーシス抑制タンパク質は、2つのメカニズムによりアポトーシスを抑制性に制御すると考えられている。1つは、BAX/BAKに結合しそのオリゴマー化を阻害するメカニズム、もう1つは、BH3-onlyタンパクに結合しその機能を阻害するメカニズムである。又、何故、BCL-2タンパク質が細胞の生と死を決めるアポトーシスを促進したり抑制したりする二つの全く相反する機能があるのは不思議に思いませんか?これに対する答えは
がん細胞にもアンテナがあり、T細胞のアンテナに結合して、「異物を攻撃するな」という命令を送ります。 すると、T細胞にブレーキがかかり、がん細胞は排除されなくなります。 このように、T細胞にブレーキがかかる仕組みを「免疫チェックポイント」といいます。
細胞周期チェックポイントとは、細胞が正しく細胞周期を進行させているかどうかを監視(チェック)し、異常や不具合がある場合には細胞周期進行を停止(もしくは減速)させる制御機構のことである。細胞自体がこの制御機構を備えている。一回の細胞分裂の周期の中に、複数のチェックポイントが存在することが知られており、これまでにG1/Sチェックポイント、S期チェックポイント、G2/Mチェックポイント、M期チェックポイントの4つが比較的よく解析されている。この機構は正確な遺伝情報を娘細胞、ひいては子孫に伝達するための、生命にとって根源的な役割を果たしていると考えられており、この機構の異常はヒトなどのがん発生の主要な原因のひとつといわれる。
PD-1/PD-L1 も免疫チェックポイント・タンパク質です。PD-1(programmed cell death – 1)受容体(別名:CD279)は活性化 T 細胞の表面に発現します。一方 PD-1 のリガンドである PD-L1(別名:B7-H1、CD274)および PD-L2(別名:B7-DC、CD273)は、通常抗原提示細胞の表面上に発現します。PD-1、PD-L1 および PD-L2 は、T 細胞応答を抑制もしくは停止させる共同抑制因子として働く、免疫チェックポイント・タンパク質です。腫瘍細胞に対する免疫システムは PD-1 と PD-L1 の反応によって必要な場合のみ発動し、自己免疫疾患となることを防ぎます。
このようにチェックポイントの意味は「阻止する」という意味で用いられすべて癌に関わっています。癌の原因はヘルペスですからヘルペスを免疫は殺しきれないないのことを知っているのでヘルペスに対しては「免疫寛容」を起こしてきょうぞんするためにがんをころしても意味がないと判断して免疫の働きも細胞周期の進行もチェックポイント(阻止する)してしまうのです。
やっとMHC 制限とかMHC 拘束の議論に到達しました。T 細胞の MHC 制限(拘束)をテストするプロセスは、通常、正の選択と呼ばれますがT 細胞の MHC 制限をテストするプロセスは存在しないのです。ここでの「検査官」は胸腺の皮質領域にある上皮細胞であり、皮質胸腺上皮細胞 (cTEC) が T 細胞に尋ねる質問は、「私が表面に発現している自己 MHC 分子の 1 つを認識する受容体はありますか?」です。正しい答えは「はい、あります!」です。なぜなら、TCR がこれらの自己 MHC 分子のいずれも認識しない場合、T 細胞は死滅するからです。自己面疾患はあり得ない病気ですからこのようなつまらない質問はする必要がないのですが病原体の非自己の蛋白質・ペプチドを認識するためには正常なT細胞として自己 のMHC 分子を持ちかつ認識するテストに合格しなければ胸腺から出ていくことはできません。従ってこのテストは本来自己免疫疾患を起こさないためのテストではないのです。
皮質胸腺上皮細胞の表面にある MHC 分子には実際にはすでに自己のペプチドが載せられているため、TCR が実際に認識するのは、自己 MHC 分子とそれに関連するペプチドの組み合わせです。cTEC のクラス I MHC 分子によって提示されるペプチドは、細胞内で作られているタンパク質のサンプルを表します。これが通常のクラス I MHC 分子の 提示です。又、皮質胸腺上皮細胞は、クラス II MHC 分子も使用して、胸腺内の外部の環境から取り込んだタンパク質の断片を提示します。これが通常のクラス II MHC 提示です。しかし、免疫学者は最近、胸腺皮質上皮細胞がクラス II MHC 分子を使用して、これらの細胞の外部から来ない多くのペプチドを提示していることを発見しました。これは、いわゆる「異常な」クラス II MHC 提示ですといわれますが、間違いです。というのは胸腺内抗原提示細胞は、主として胸腺皮質に存在する上皮細胞であるのでクラス II MHCを持っていても「異常な」クラス II MHC 提示ではないのです。
細胞は、飢餓の時期、つまり細胞成分の合成に必要な原材料が限られている状況に対処するのに役立ついくつかのメカニズムを進化させてきました。そのような生存ツールの 1 つが、オートファジー (文字通り「自己食」) と呼ばれるプロセスです。細胞が飢餓状態になると、細胞質の一部を膜で囲み、それがリソソームと融合します。その後、オートファジーとは細胞質成分 (タンパク質など) はリソソーム酵素によって分解され、再利用できるようになります。驚くべきことに、胸腺皮質上皮細胞は、オートファジーを使用して自身の細胞内タンパク質を捕捉し、短いペプチドに消化し、クラス II MHC 分子を使用して表面に提示することもできます。オートファジーを利用してこの異常なディスプレイ(提示)を準備することで、胸腺皮質上皮細胞は胸腺内の T 細胞に提示できる自己ペプチドの範囲を大幅に拡大するのではなくて元々胸腺皮質上皮細胞は胸腺内に住んでいる抗原提示細胞であるからです。おそらく、これにより T 細胞がクラス II MHC 分子と結合できるペプチドの組み合わせを認識し、生存のために積極的に選択される可能性が当然、高くなるのは抗原提示細胞であるからです。例のごとく「おそらく」とか「可能性」というあいまいな言葉が使われて推測で終わっているのは、この世に存在しない自己免疫疾患という病気を捏造するために免疫学者の牽強付会の努力に乾杯しましょう。ワッハッハー!!!オートファジー (Autophagy) とは何でしょうか?細胞が持っている、細胞内のタンパク質を分解するための仕組みの一つである。酵母からヒトに至るまでの真核生物に見られる機構であり、細胞内での異常なタンパク質の蓄積を防いだり、過剰にタンパク質合成したときや栄養環境が悪化したときにタンパク質のリサイクルを行ったり、細胞質内に侵入した病原微生物を排除したりすることで生体の恒常性維持に関与している働きなのです。いわば緊急事態で仕方なく発揮される働きがオートファジーなのです。ましてや恒常的に自己免疫疾患を起こさないように行使されるものではないのです。
自己免疫論者の唱えるMHC 制限の論理矛盾を見てみましょう。試験の合間に少し立ち止って、重要な質問をしてみましょう。なぜ T 細胞は、自己 MHC 分子によって提示されるペプチドを認識できるかどうかを確認するためにテストする必要があるのでしょうか。結局のところ、ほとんどの人間は一生を「外来」の MHC 分子 (移植された臓器など) を見ることなく終えるので、MHC 制限は自分の MHC 分子と自分の MHC 分子を区別することではありません。いいえ、MHC 制限は外来と自己とは関係ありません。焦点が重要なのです。講義 4 で説明したように、T 細胞が MHC 分子によって提示される抗原に焦点を合わせるようにシステムを設定する必要があります。B 細胞の受容体と同様に、T 細胞の受容体は遺伝子セグメントを混合して組み合わせることで作成されるため、非常に多様です。その結果、T 細胞で発現される TCR のコレクション(集合体)には、B 細胞の受容体と同様に、提示されていない抗原を認識するものが多数あることは間違いありません。これらの T 細胞は排除する必要があります。そうでなければ、MHC 分子による抗原提示の素晴らしいシステムは機能しません。したがって、正の選択 (MHC 制限) が非常に重要なのは、すべての成熟 T 細胞が自己 MHC 分子によって提示された抗原を認識する TCR を持つシステムを構築するためです。後で負の選択も出てきますが後でわかりますがここで使われる正も負も科学の一分野である医学という学問分野で用いるべきではないのです。
自己寛容性のための胸腺テストも全く嘘ですが自己免疫論者の論理がどこが矛盾があるかを指摘しましょう。因みにcoreceptorとはなんでしょうか?coreceptorの訳は 「補助受容体、共受容体、共役受容体、共同受容体、共働レセプター、コレセプター」などといくつかありますが最も的確な訳はcoreceptorとは「補助受容体」です。 CD4とCD8は典型的なコレセプターであるのはT細胞Receptor(TCR)を補助できるレセプターがCD4とCD8であるからです。
胸腺皮質で正の選択(MHC 制限)が行われている間、または少し後に、T 細胞は CD4 または CD8 のいずれかの補助受容体分子の発現を停止します。予想どおり、これらの細胞はその後、CD4 または CD8のどちらかを発現するので単一陽性 (SP) 細胞と呼ばれます。ところがT 細胞が CD4 または CD8 補助どちらかの受容体の発現を「選択」する正確なメカニズムは今なお不明です。長い間探求されていますがまだ答えは出ていません。ただし、補助受容体の選択は、特定の T 細胞が皮質胸腺上皮細胞上のクラス I またはクラス II MHC 分子によって表示される同族抗原(cognate antigen) を認識するかどうかによって決まるという見方が浮上していますがこれも怪しい推測にすぎません。しかしこの論理も疑問があります。何故ならばまず第一に同族抗原(cognate antigen)とは特定のT 細胞が認識できる抗原ですから補助受容体の選択は同族抗原(cognate antigen) を認識するかどうかによって決まるわけですからこの段階ではT 細胞は同族抗原(cognate antigen) を認識できるほど成熟していないので同族抗原(cognate antigen) を認識できるはずがありません。更に特定の T 細胞はどんなT細胞なのかの説明する必要があるのに特定の意味の説明がありません。いずれにしてもこの特定の T 細胞はまだ未熟でTCR(T細胞受容体)も持っていないのですよ。
更に、皮質胸腺上皮細胞はAPCではないのでクラス II MHC 分子を持っているわけはないでしょう。たとえば、T 細胞の受容体が皮質胸腺上皮細胞に発現しているクラス I MHC 分子によって表示される抗原を認識すると、T 細胞表面の CD8 補助受容体が「パーティーに参加」し、MHC 分子にクリップし結びつくことはありうるでしょう。これが起こると、その T 細胞上の CD4 分子の発現がダウンレギュレーション(低下)しますと書かれていますが何故低下するのかは説明されていません。同様に、受容体が皮質胸腺上皮細胞が持っていないクラス II 分子によって表示されるペプチドを認識する T 細胞は CD4 T 細胞になり、その細胞上の CD8 補助受容体の発現はオフになりますと論じていますがその根拠がありません。この戦略が機能するのは、CD8 補助受容体がクラス I MHC 分子にのみ結合し、CD4 共受容体がクラス II MHC 分子にのみ結合するためですが証拠がないので仮定の話にすぎません。
TCR が自己 MHC に結合したペプチドの複合体を嘘で認識する幸運な T 細胞は、その表面に CCR7 ケモカイン受容体を発現し始め、胸腺皮質から髄質と呼ばれる胸腺の中心領域に進みます。髄質胸腺上皮細胞には CCR7 のリガンドが豊富にあります。2 番目のテスト、つまり「自己の成分に対してT 細胞が反応できないように寛容がおこるテスト」が実施されるのは胸腺髄質です。この検査は、寛容をおこさないT 細胞を排除するための検査なのでネガティブ セレクション(負の選択)と呼ばれることがよくあります。CCR7やCCR7 のリガンドとは何でしょうか?CCケモカイン受容体7型(CCR7受容体)は、ヒトではCCR7遺伝子によってコードされるタンパク質である。この受容体には、ケモカイン(CCモチーフ)リガンド19(CCL19 /ELC)と(CCモチーフ)リガンド21(CCL21)の2つのリガンドが同定されている。受容体は免疫細胞をリンパ節などの免疫器官に誘導します。これは、胸腺のT細胞の発生成熟に重要です。CCR7受容体は、主に高等免疫である適応免疫細胞タイプである胸腺細胞、ナイーブT細胞とB細胞、制御性T細胞、中心記憶リンパ球以外の自然免疫の樹状細胞に発現しています。自然免疫の樹状細胞に発現して樹状細胞の成熟を刺激する働きがあります。CCR7受容体は自然免疫の樹状細胞に発現していることはCCR7受容体は自己免疫疾患にならない証拠となります。CCR7は、リンパ節や脾臓などのさまざまな二次リンパ器官へのT細胞のホーミングや、脾臓内でのT細胞の輸送にも関与しており外来から人体に侵入したヘルペスをはじめとする非自己の病原体がたむろしている部位にCCR7はT細胞(ヘルパーT細胞とキラーT細胞)を送る仕事をしているのです。T細胞のホーミングとは?免疫系の主役となるTリンパ球は,血管系とリンパ系を絶えず循環し,外界からの細菌やウイルスなどの異物侵入に対してパトロール役として機能する。骨髄や胸腺などの一次リンパ組織で産生されたTリンパ球は,二次リンパ組織であるリンパ節に流入し,さらにそのリンパ節を離れて全身を巡り,再び同じ特定の二次リンパ組織に戻ってくる. この現象をリンパ球ホーミングという.
ネガティブ セレクション(負の選択)中に T 細胞に尋ねられる検査の質問は、「私の表面にある MHC 分子によって表示される自己ペプチドを認識しますか?」です。正解は「ありえない!」です。まずこんな愚かな質問を恥ずかしげもなく考え出す学者がいかに馬鹿かわかりますか?自分のpeptide(ペプチド)を認識しないT細胞がありますか?自分のpeptide(ペプチド)蛋白を認識できるからこそMHCが全く異なる他人の移植臓器を拒絶してしまうことを知らないバカの極みというべき医者なのです。すべてのT細胞は自己自身のペプチド抗原が乗せられた自己 MHC複合体の二つを認識しても活性化しないのは自己自身のペプチド抗原には反応しないからです。何度も言いますが自己自身のペプチド抗原は絶対に危険信号(danger signal)にはならないのです。つまり自己のpeptideを認識することとT細胞が活性化されて自己免疫疾患を起こして戦いが始まり炎症を起こすこととは全く別の事柄であることを世界中の賢い免疫学者は知らないのです。「私の表面にある MHC 分子によって表示される自己ペプチドを認識しますか?」という質問は自己ペプチドを認識したT細胞は活性化して自己免疫疾患が起こりますという問題と自己のペプチドを認識することとは全く別問題であることを、自己免疫論者は全く理解していないバカなのです。
自分自身の抗原に反応する可能性のある T 細胞を排除するこの 2 番目の検査は非常に重要です。実際、このような自己反応性 T 細胞が削除されなければ、自己免疫疾患が発生する可能性があります。かれらは相変わらず「可能性があります」にこだわっていますがいずれにしても「可能性はゼロなのです。」何故ならば、言うまでもなく自然免疫のレベルで自分のpeptideを認識してもしなくても自己反応性 T 細胞があるとしても自分のペプチドは敵とみなすことはないので自己免疫疾患が発生する可能性はゼロです。何故ならば自然免疫が病原体の非自己だけのペプチドだけを敵と認識するだけで,高等免疫のT細胞にMHC 分子ⅠもMHC 分子Ⅱも持っている活性化した大食細胞と樹状細胞とBリンパ球などのAPCは危険信号(danger signal)として提示しないのです。従ってAPCのMHC 分子に乗せられない自己のペプチドにはT細胞やB細胞の高等免疫は絶対に免疫応答が自分のペプチドを異物と認識させてもらえないので免疫応答が起こることはないのです。従って自己免疫疾患は絶対にあり得ないので「自己寛容」や「MHC拘束」を論じることほど虚しいことは他に何もないのです。お金と時間の無駄そのものです。残念です。
自己のペプチドは認識できてもできなくても自己抗原を認識する Th 細胞は、B細胞に抗体を作らせるつらせる手助けをしても作成し、自分自身の分子 (血液中のインスリン タンパク質など) を破壊対象としてタグ付けするのを助ける可能性がありますと阿呆な免疫論者は言い続けますが大間違いです。または、自分自身の細胞を攻撃する CTL が生成される可能性がありますとも医学者は言いますがあらゆる10万個以上もあるMHCには自己のpeptideが満載されて乗せられていますが自己反応性のT細胞は存在しないので自己免疫疾患は絶対にありえないのです。
ここから髄質胸腺上皮細胞(mTEC)の働きについての間違った自己免疫論の間違った説明が始まります。自己免疫学者の間違った言い分を聞きながら批判していきましょう。
自己抗原に対する寛容をスクリーニング(調査)する細胞には 2 つのタイプがあり、どちらの細胞タイプも、T 細胞の MHC拘束の有無 (正の選択) をテストした皮質胸腺上皮細胞(cortical thymic epithelial cellryakusite略して cTEC)とは異なります。T 細胞の自己寛容をテストする細胞タイプの 1 つが髄質胸腺上皮細胞 (medullary thymic epithelial cell略してmTEC) です。これらの細胞は、MHC拘束をテストする皮質胸腺上皮細胞の親戚であり、特に「寛容テスター」として適している 2 つの特性があります。
MHC拘束とはもっと具体的には何でしょうか?MHCは、細胞内外のタンパク質が細胞内でペプチドに分解されたMHCとペプチドの複合体になって初めて細胞表面に輸送してT細胞に抗原として提示することで、T細胞が活性化して免疫応答が開始されるのです。病原体を免疫で処理するための病原体の持ちペプチドだけを認識するだけでなく自分のMHCと合体して複合体になったペプチドしか敵だと認識しないのを「ペプチドはMHCに拘束される(MHC拘束)と表現するのです。
皮質胸腺上皮細胞と同様に、髄質胸腺上皮細胞(mTEC) はオートファジーを使用して自身の「内部」のタンパク質をペプチドに消化し、これらのペプチドをクラス II MHC 分子で提示します。クラス II MHC 分子はAPC(抗原提示細胞)しかもっていないのですが
細胞)内で生成されたタンパク質がクラス II MHC 分子によって表示されるこの規則を破った提示は、負の選択中にほとんどの自己反応性ヘルパー T 細胞を排除するために使用できる多様な自己抗原源を提供します。
ただし、まだ問題があります。すべての細胞が生成する「共通」タンパク質に加えて、「組織特異性」のタンパク質が多数(推定では数千)あります。これらの組織特異性タンパク質は、各臓器または組織タイプにその独自性を与えるものです。たとえば、心臓を構成する細胞によって生成されるタンパク質には、その臓器に固有のものがあります。また、腎臓細胞によって生成されるタンパク質には、腎臓特異性があります。したがって、胸腺での耐性試験(自己免疫寛容)を完全なものにするには、学生である T 細胞を検査する「材料」に組織特異性タンパク質を含める必要があります。そうしないと、キラー T 細胞が胸腺を離れるときに、その一部が耐性のない(自己免疫寛容のない)組織特異性タンパク質に遭遇し、肝臓、心臓、腎臓を破壊し始めることになります。これはよくありません。
胸腺の皮質と髄質の違いは何ですか?胸腺組織は免疫応答の司令塔であるT細胞の産生に必須の中枢リンパ組織であり,髄質および皮質からなります。 皮質はおもに自己のMHC分子への反応性の獲得および幅広い抗原に反応しうるレパトア(抗原認識の多様性)の形成を担い,髄質は中枢性の自己寛容の成立において中心的な役割をはたすといわれますがウソです。胸腺組織は骨髄で生まれた裸の赤ちゃんであるT細胞を衣類を身に着けた完璧な成人の大人にするために存在しているのです。この意味をゆっくり解説していきます。B細胞も免疫寛容のための複雑なシステムは不必要なのです。進化論的に自己のタンパク質やペプチドを高等免疫(獲得免疫)であるB細胞もT細胞は自己成分なので敵とみなす免疫の働きは一切存在しないように進化しました。それはどのようにしてかというとB細胞もT細胞も自己でない異物を攻撃するために生まれた強い戦士ですから自分の味方である自己を攻撃すると軟化の間違いが生じて自己を攻撃しないように自然免疫の三つのAPCに非自己の敵だけを教えるように進化したのです。自己を攻撃しないように二つの保証を付けたのです。一つ目はAPCが敵として認識するのは非自己の病原体だけであり二つ目はT細胞も敵として非自己成分だけを認識するであり、逆の言い方をするとAPCが敵として認識できないのは自己の成分だけであり同時にT細胞も敵としては自己成分は認識しないのです。実際には自然免疫である抗原提示細胞だけが非自己である微生物のペプチド蛋白分子を敵と認識してその敵をT細胞も敵として認識して初めて獲得免疫が開始されるのです。言い換えると絶対間違いを犯さない「自然免疫」の指導通りに「獲得免疫」も同じ敵を攻撃し始めるのです。自然免疫の活性化された樹枝状細胞は獲得免疫の正に忠実な橋渡し役を務めているのです。
幸いなことに、髄質胸腺上皮細胞(mTEC)は、多くの組織特異性抗原の発現を促進する AIRE(auto‐immune regulator訳して「自己免疫制御蛋白質」略してAIRE) と呼ばれる転写因子を生成します。AIRE(auto‐immune regulator訳して「自己免疫制御蛋白質」略してAIRE)とは何でしょうか? Aire遺伝子は自己免疫を制御するタンパク質と訳されているように自己免疫疾患にならないためには必須の遺伝子。 主に胸腺に存在する胸腺髄質上皮細胞に発現し,組織特異的抗原の発現を促すことで自己反応性T細胞を除去する。Aire遺伝子の変異により発症する臓器特異的自己免疫疾患と皮膚粘膜カンジダ症を主な症状とする常染色体劣性疾患といわれます。残念ながらAire遺伝子によって発現する転写因子によってどのような遺伝子がどのようなタンパク質に発現されるのかについては一切触れられてはいないのです。とにかくこの世に存在しない自己免疫疾患をゴリ押しでお金儲けのために医薬業界が一致団結してウソをつきまくっているのです。この犯罪は医薬業界全体の組織犯罪なのです。その虚偽の犯罪の結果、髄質胸腺上皮細胞は、通常の共通タンパク質に加えて、1,000 を超える組織特異性タンパク質を発現しますと言いふらしているのですが、1,000 を超える組織特異性タンパク質を発現するメカニズムは一切触れられていません。たったひとつの髄質胸腺上皮細胞が発現するAire遺伝子のよって発現する転写因子によって1,000 を超える組織特異性タンパク質が発現されるということはありうると思いますか?
いずれにしろ、組織特異的抗原に対する自己寛容の問題については、まだ不確実な点がありすぎるとまで自己免疫論者も認めています。たとえば、mTEC(髄質胸腺上皮細胞) が体内にある組織特異的タンパク質のすべてを発現するのか、それとも大部分だけを発現するのかはわかっていませんとまで述べているのですがやはり自己免疫疾患があるといい続けなければ医薬業界は即、破綻してしまうので自己免疫疾患はないといえないのです。
嘘をついて治りもしない免疫を抑える薬を作って病気は治らずヘルペスを増やし続けて、逆に自己免疫疾患といわれる症状を生み出しているヘルペスを無視して患者を苦しめて最後は癌を作って金を儲けるアコギな資本主義医薬業界の繁栄は資本主義がつぶれない限り永遠に続きます。悲しいですね。お金で買えない快楽は「健康」だけなのですがすべてにおいて真実に無知な無批判な大衆は医薬業界に騙され続けるのが特権ですね。死ぬのが仏の世界です。
胸腺樹状細胞(TDC)とはなんでしょうか?胸腺における自己抗原に対する寛容性の検査に関係する 2 番目の細胞タイプが胸腺樹状細胞 (TDC) ですといわれていますが、間違いです。胸腺 DC(thymic dendritic cell,略してTDC、訳して胸腺樹枝状細胞)は樹枝状細胞ですから特徴的なヒトデのような形をしていますが、自然免疫の抗原提示細胞(APC)である「遊走性」樹状細胞とは異なります。髄質 TDC(medullary thymic dendritic cell,略してmTDC)は胸腺髄質の「常在細胞」であり、骨髄由来の前駆細胞から髄質で発生します。TDC の興味深い点は、胸腺環境から通常の方法で抗原を獲得することに加えて、それらが提示する抗原の一部が mTEC によって「与えられる」というよりも提示されます。実際、mTEC からの 自己のMHC+自己ペプチド複合体は、何らかの方法で胸腺樹状細胞に「引き渡され」、CD4+ 細胞と CD8+ 細胞の自己寛容性の検査に使用されるようですと言われますが間違いです。というのは免疫細胞から別の免疫細胞に「物質である蛋白質」をちぎって与えることは絶対に不可能であるからです。正しくは胸腺は血液が豊富ですから血液で運ばれてきた自己のペプチドを取り込んで自己のMHCに乗せて複合体を形成しただけなのです。この複合体を胸腺で成熟し切ったT細胞に見せつけてもT細胞は反応することは無いことを確かめるためです。にもかかわらず自己免疫論者はこの引き渡しがどのように行われるか、そしてなぜそれが重要なのかは謎のままですというのは当たり前なのです。何故ならばすでに証明したように自己免疫疾患は存在するわけはないので胸腺で「正の選択」とか「負の選択」などにかかわる現象は一切存在しないからなのです。明らかに、とりわけ胸腺における負の選択については、まだ多くのことが解明されていないのですとはっきり述べているのにもかかわらず、この本を書いた世界でウソつきの一流の免疫学者自身が書き記しているのにもかかわらずしかも自己免疫疾患は何の証拠もないのに主張しているのにあきれ返ります。自己免疫疾患が発表されて80年近くたっているのに今でも何の進展もない理論であるのに何故自分の誤りに気が付かないのでしょうか?
先進国ではワクチンと抗生物質ができてから病気つまり感染症で死ぬ人はほとんど皆無になりました。だからこそ新たに病気で死ぬ「ウソの疾患」を作り出してたのです。その代表が自己免疫疾患です。病気がなくなれば医薬業界が崩壊することを恐れているのです。病気がなくなれば医者冥利に尽きる話でしょうに。自己免疫疾患が存在すると主張するすべての論文は嘘です。ただ一つ自己免疫疾患関する真実の論文は松本仁幸医学博士の「自己免疫疾患はない。自然免疫のAPCが自己の成分を敵と認識しないからありえないのです。自己免疫疾患が本当に存在する病気であればあれば人類は誕生していないしとっくの昔に滅び去っているのです。ワッハッハ!!!!!!!!すべての人の胸腺は20歳すぎると急激に萎縮します。胸腺は体の臓器の中で最も早く老化する臓器で、成人では小さくなって脂肪組織に置き換わってしまうので自己免疫疾患で成人は死んでいくはずなのに最後は癌で死んでいくのは何故でしょうか?自己免疫疾患は存在しない「偽の病気」であるからです。この事実だけでも「自己免疫疾患は絶対にないと証拠になるのに松本博士だけしか知っているのは何故でしょうか?皆さん考えてください??????
胸腺におけるややこしい「正の選択テスト」とあり得ない「負の選択のテスト」を極めて簡単に説明しましょう。胸腺におけるT細胞の成熟は、「正の選択」と起こらない「負の選択」という2段階で行われる。 骨髄から中枢リンパ組織である胸腺に血液から移入してきたT細胞の前駆体は、胸腺で分化・成熟する過程で「正の選択」により正常な異物を認識できる能力を獲得し、一方起こっていない「負の選択」により自己を異物と認識しない能力を獲得する。 この「負の選択」の過程が自己成分に対する中枢性免疫寛容であるといわれますがこの「負の選択」の過程が自己成分に対する中枢性免疫寛容であるのですがこれは嘘です。
胸腺におけるこの虚偽の最終結果は、胸腺皮質上皮細胞によって提示される自己 MHC とペプチド複合体を認識する受容体を持つ T 細胞の集合体であるといわれますが集合体どころか自己 MHC ペプチド複合体を認識するT細胞は一個すらもないのです。が、胸腺樹状細胞または髄質胸腺上皮細胞上の MHC 分子によって提示される自己抗原は認識しないのは、もともと自然免疫のあらゆる細胞も獲得免疫といわれる高等免疫のT細胞も自己のペプチドや分子を敵と認識出来るわけはないし、認識する必要もないのです。脊椎動物や人類が誕生したとき以来自己免疫疾患があれば人類は滅びています。胸腺は獲得免疫だけのT細胞を完璧に行うための教育を行うだけのT細胞の成長・成熟の場に過ぎないのです。人体の敵となる非自己の病原体だけを認識して攻撃するために生まれたのがT細胞なのです。
そもそも「免疫」とは、人間にとって「異物」となる細菌やウイルスなどの微生物、病原体とよばれる自分の体には元々ないもの(非自己)から、自分たちの体(自己)を守るしくみ炉して誕生したのです。「免疫機能」のことを「生体防御機能」ともいいます。これが免疫の絶対的な定義です。
だのにいつから自分たちの体(自己)を攻撃するようになったのでしょうか?漫画の世界ですね。ワッハッハ!!!!!!!!免疫機能は加齢とともに低下しますが、自然免疫系よりも獲得免疫系の方が著しいのです。獲得免疫の能力は、20代頃がピークであり、40代ではその半分に低下するのです。胸腺が老化するのはなぜでしょうか?この胸腺の老 化による萎縮は 2 つの原因で発生するのです。 1 つは骨髄から 胸腺に流入する T 前駆細胞の産生が骨髄の老化による変化で減少することであり,もう1 つは T 細胞分化の場でナーシング細胞として働く胸腺上皮細胞の老化による機能低下です。ナーシング細胞とは何でしょうか?胸腺ナース細胞とも呼ばれ骨髄から血管によって運ばれてきた多数の未熟なリンパ球を包み込んで球形の形態をとっている胸腺上皮細胞です。未熟なリンパ球を保育するように見えるからナース細胞と呼ばれるのです。
胸腺は成長とともに小さくなりますか?
胸腺は縦隔の上(上縦隔)かつ前(前縦隔)の方で、胸骨の裏側にあります。 大きさは成長とともに変化します。 誕生時には10グラムから15グラム、思春期には最大の30グラムから40グラムになります、その後は徐々に小さくなり、成人以後は、脂肪の一部のようにみえます。胸腺は放射線や副腎皮質ホルモンなどに曝露されると萎縮するが特にT細胞を盛んに産出している時期は感受性が高い。 胸腺中のリンパ球が最も多いのは思春期(10代前後)でピーク時の胸腺は30 – 40gに達する。 その後は急速に萎縮し脂肪組織に置き換わる。 この胸腺の退縮は70歳までにほぼ完了してしまう胸腺の仕事は廃絶します。
胸腺が委縮しはじめさらに脂肪組織に置き換わると胸腺の機能が低下して新たなる問題が出てきます。元々自己免疫疾患は存在しないので胸腺の働きがなくなっても全く問題はないのですが一つだけ難問が突然出現することになります。本来胸腺の機能は骨髄の造血幹細胞から生まれた毎日、約 6,000 万個の二重陽性細胞のT細胞の前駆細胞を分化・成熟させて完璧なヘルパーT細胞やキラーT細胞に育て上げることですが胸腺が脂肪組織に変わってしまうと新しい成熟し分化したヘルパーT細胞やキラーT細胞が胸腺から生まれなくなります。免疫の司令官でありかつ獲得免疫の中枢であるT細胞が生まれなければどのようにして病原体から人体を防御できるのでしょうか?その答えは「自己免疫疾患は存在しない」の論文を書き終わってから述べます。乞うご期待!!
これらの意味のないテストに合格した「胸腺卒業生」は、表面に高レベルの多くの分子で構成された T 細胞受容体と、CD4 または CD8 共受容体(補助受容体)のいずれか一つだけを発現します。若者の胸腺では毎日、約 6,000 万個の二重陽性細胞がテストされますが、胸腺から排出されるT 細胞受容体とCD4 または CD8 共受容体(補助受容体)のいずれか一つだけを発現できる単一陽性細胞は約 200 万個のみのT細胞は胸腺から出ていきます。しかし胸腺から出ていくこの選ばれた200 万個のテストに合格したT 細胞はあくまでも「正の選択(MHC 拘束)」と「負の選択(寛容誘導)」の自己免疫疾患を起こさないテストに合格したわけではないのです。何故ならば答えがまるっきり反対の試験に合格できるわけはないでしょう。こんな試験がこの世に存在するのは自己免疫疾患が存在しないので「正の選択(MHC 拘束)」と「負の選択(寛容誘導)」の正しいテストはあり得ないからです。無理やり間違った論理を立てて金儲けをしようとしたから矛盾だらけなのです。残念です。試験に合格できなかった生き残ったT 細胞は役に立たないと判定されるのは正の選択(MHC 拘束)」と「負の選択(寛容誘導)」のテストに落ちたからではなくT 細胞受容体とCD4 または CD8 共受容体(補助受容体)のいずれか一つだけを発現できることができなかったT 細胞だけがアポトーシスによって死滅し、胸腺内のマクロファージによってすぐに食べられてしまうのは当然なのです。
免疫学者は正常なT細胞が胸腺の血管からどのように排出されるのかまだわかっていませんが、血液を介して皮質髄質接合部付近から排出されると考えています。何回も念を押しますが正常なT細胞というのは自己のMHCを認識しないT細胞では無いのですよ。胸腺のみならずT細胞も一切自己免疫疾患には関りがないのです。自己免疫疾患は医学者が捏造したお金が儲かる病名にすぎないのです。正常なT細胞は病原体の異物と自己のMHCの二つを同時に認識して初めて「敵を認識する」という仕事を行っているのですよ。
永遠に解けないMHC 拘束と寛容誘導の謎について述べます。「MHC 拘束と寛容誘導の謎」が解けないのは「自己免疫疾患は存在しない病気」である証拠なのです。さて、これまでの間違だらけの論文を注意深く私と一緒に勉強してきた方なら、T 細胞がどのようにして両方の試験に合格できるのか疑問に思うかもしれませんどころか全て「ウソ」であることに気が付かれていることでしょう。何故ならば結局のところ、MHC 制限の試験に合格するには、TCR が MHC と自己ペプチドを認識しなければなりません。しかも、自己の成分に対して寛容である試験に合格するには、TCR が MHC と自己ペプチドを認識できない必要があります。こんな矛盾に満ちた自己免疫疾患の理論が大手を振って世界中の医薬業界をお金儲けのためだけに支配しているのですよ。2 つの試験が互いに打ち消し合って、T 細胞が絶対に合格できないのです。これがウソ100%の自己寛容の謎の本質ですから、謎というよりもウソ偽りの本質というべきなのです。
T 細胞受容体が敵のペプチドに連結することによって、どのようにして正の選択 (MHC 制限) と負の選択 (寛容誘導) の両方が生じるのでしょうか? 実際、それはそれよりもさらに複雑です。なぜなら、T 細胞が胸腺で両方の試験に合格して胸腺を出て侵入者と出会ったときに、そのT 細胞の TCR はAPC(抗原提示細胞)が自己 MHC 分子によって提示された侵入者由来のペプチドに遭遇したときに、活性化されたシグナルをT 細胞の核に送ることが出来なければならないからです。ややこしいでしょう。難解でしょう。だから免疫はすごく面白いのです。そのT細胞がAPCによって活性化され敵を認識したという信号を核の遺伝子の送る話をしましょう。
「ゴルディロックス」仮説とは何でしょうか? まず「ゴルディロックス」という英語の本来の意味は「金髪のきれいな娘」です。「ゴルディロックス」仮説は「ゴルディロックスの原理」とも言われます。ゴルディロックスの原理は本来、経済学用語です。 『三匹の熊』の童話の喩えを借りて名付けられたものです。物語の中にゴルディロックスという名前の少女が登場し、三種のお粥を味見したところ、熱すぎるのも冷たすぎるのも嫌で、ちょうど自分の口に合うよい親和性の温度のものを選ぶという話から生まれた原理なのです。
MHC 制限と寛容誘導につながるイベントは、T 細胞の活性化に関係するイベント、つまり細胞間接着、TCR クラスター形成、および共刺激と類似していることが知られています。しかし、免疫学者を悩ませている疑問は、同じ TCR が MHC ペプチド複合体と関わるときに、どのようにして 3 つの非常に異なる結果 (正の選択、負の選択、または活性化) をシグナルするかということです。残念ながら、私はどのようにして 3 つの非常に異なる結果 (正の選択、負の選択、またはT細胞の活性化) をシグナルするかというこの疑問に答えることができませんとこの論文を書いた著名な自己免疫論者は泣き言を言っています。もし、この問いに答えることができれば、ノーベル賞を受け取るためにスウェーデンに向かうことになりますと冗談を飛ばしていますがウソの原理ですから絶対にノーベル賞は無理です。しかし、免疫学者は「親和性モデル」、つまりゴルディロックス仮説とも言えるものを支持しています。この仮説は、胸腺での正の選択と負の選択の両方を生き残るには、T 細胞が「ちょうど良い」受容体を持っている必要があると述べています。「ちょうど良い」受容体とは何でしょうか?受容体は気分で動くものではないでしょうに???実際、胸腺では、T 細胞の正の選択 (生存) は、TCR と皮質胸腺上皮細胞に提示された MHC 自己ペプチドとの間の比較的弱い相互作用 (TCR が提示された抗原に集中することを保証するのに十分な強さの相互作用) によって生じると仮定されています。この曖昧な仮定も医学に応用するのは間違っています。医学は冷厳たる事実に基づくものです。人の命を扱う医学はいつまでもいつまでも仮定の世界に安住すべき学問ではないのです。次に、TCR と髄質胸腺上皮細胞または胸腺樹状細胞に発現された MHC 自己ペプチドとの間の相互作用が強すぎてはならず、そうでないと細胞死 (負の選択) が生じます。そして最後に、T 細胞が胸腺を離れた後、その TCR と専門の抗原提示細胞によって提示された MHC ペプチドとの間の相互作用が、活性化を引き起こすのに十分な強さでなければなりません。十分な強さとは何で決めるのですか???
もちろん、固定されたTCR は 3 つの状況すべてで変化しない同じタンパク質なので、MHC ペプチドと T 細胞受容体のこれらの 3 つの相互作用の効果を、生、死、または活性化のどれにこれほどまでに異なるものにするのかが疑問です。と論者自身が認めています。1 つの重要な要素は、信号を「送信」する細胞の特性であると思われます。MHC 制限の場合、これは皮質胸腺上皮細胞です。寛容誘導の場合、細胞は骨髄由来樹状細胞または髄質胸腺上皮細胞です。活性化の場合、送信者は特殊な抗原提示細胞です。これらの送信細胞は非常に異なります。たとえば、皮質胸腺上皮細胞のプロテアソーム (クラス I 表示用のペプチドを作成するためにタンパク質を切り刻む機械) は、負の選択を担う細胞のプロテアソームとは微妙に異なります。これは、これらの検査細胞によってクラス IMHC 分子にどの自己ペプチドが提示されるかに影響する可能性があります。さらに、胸腺皮質上皮細胞がクラスⅡMHC分子上に提示するためのペプチドを準備するために使用する酵素の一部は、胸腺髄質1aの検査細胞が使用する対応する酵素とは異なります。この論理は自己免疫論者自身が自己免疫疾患はないと告白しているのと同然です。
また、さまざまな送信細胞は、それらが発現する細胞接着分子と、それらが表面に表示するMHCペプチド複合体の数またはタイプが異なる可能性があります。このような違いは、T細胞受容体を介して送信される信号の強度に劇的な影響を与える可能性があります。さらに、異なるタイプの送信細胞は、共刺激分子(補助刺激分子)の異なる混合物を発現する可能性があり、共刺激信号は、TCR-MHC-ペプチドの関与から生じる信号の意味を変える可能性があります。疑問だらけの自己免疫疾患がありえないという答えになっていますね。ワッハッハ!!!!!信号を送信する細胞が異なるだけでなく、「受信者」(T細胞)も検査ごとに変わる可能性があります。T細胞の表面にあるTCRの数は、細胞が検査されるにつれて増加することが知られており、T細胞が成熟するにつれてT細胞内の「配線」が変化する可能性もあります。 TCR 密度とシグナル処理におけるこれらの違いは、さまざまな種類の送信細胞によって生成されるシグナルの解釈に影響を与える可能性があります。
MHC 制限/寛容誘導パズルのピースの多くは発見されていますが、免疫学者はまだそれらを完全に一貫した図にまとめることができていません。さらなる研究が必要です。が研究は100%無駄です。「無い」現象を「有る」と証明することほど不可能なことは無いからです。
無知による寛容とは何でしょうか?私たち自身のタンパク質を認識できる受容体を持つ T 細胞のほとんどは胸腺で除去されますといわれていますが自己のMHCはタンパク質ですよ。この自己のMHCに乗せられた非自己のタンパク質であるペプチド複合体をT 細胞は同時に認識しているのですよ。しかし、胸腺での中枢寛容誘導は絶対確実ではありません。もしそうなら、すべての T 細胞をあらゆる可能性のある自己抗原でテストする必要がありますが、それは要求が多すぎますといわれますが自己免疫疾患はT細胞は自己抗原を認識することはできないので、要求が多いとか少ないには全く関係ないのです。
胸腺に豊富に存在する自己抗原に対する親和性が高い受容体を持つ T 細胞は、そこで削除される可能性が非常に高くなります。しかし、自己抗原に対する親和性が低い受容体を持つ T 細胞、または胸腺にほとんど存在しない自己抗原を認識する T 細胞は、負の選択を受ける可能性が低くなります。それらは中枢寛容誘導の隙間を「すり抜ける」だけかもしれません。幸いにも、この可能性に対処するためのシステムが構築されていますのも嘘です。
バージン T 細胞は二次リンパ器官を循環しますが、組織には出られません。これは間違いです。というのは二次リンパ器官を循環するバージン T 細胞はリンパ管を経由して必ず心臓に戻ります。心臓から血液に入り二次リンパ器官のリンパ節に入る毛細血管のHEVから様々な二次リンパ器官へ戻ります。が、毛細リンパ管は組織においては穴が開いたままの盲管になっていますから必ずそこで組織とつながっているからです。となっているのでの交通パターンにより、これらのバージン T 細胞は、リンパ節でAPC に遭遇して活性化される可能性が最も高い体の領域に運ばれますがこれは最初は偶然です。しかし、バージン T 細胞を組織から遠ざける移動制限も、自己寛容の維持に重要ですというのも嘘です。その理由は、通常、バージン リンパ球が活性化される二次リンパ器官に豊富に存在する自己抗原は、T 細胞が寛容化される胸腺にも豊富に存在するからです。したがって、バージン T 細胞がたどる移動パターンの結果として、二次リンパ器官の豊富な自己抗原によって活性化される可能性のあるほとんどの T 細胞は、胸腺の同じ豊富な自己抗原によってすでに排除されていることになりますといわれていますがこれも嘘です。自己抗原は存在しないからです。
逆に、胸腺では比較的まれな自己抗原を受容体が認識する T 細胞は、胸腺での削除を免れる可能性があります。これも嘘です。ただし、これらの同じ抗原は通常、二次リンパ器官で非常に低濃度で存在するため、潜在的に自己反応性の T 細胞を活性化することはありません。したがって、二次リンパ器官にはまれな自己抗原が存在し、T 細胞にはそれらを認識できる受容体がありますがこれも嘘です。これらの T 細胞は通常、機能的にそれらの存在を「知らない」ままですどころかそもそも自己抗原を認識することはできないのですべてのT細胞は死ぬまで自己抗原を「知らない」ままなのです。自己抗原があまりにもまれであるため、活性化を誘発できないためです。これも嘘です。人体は膨大な自己抗原で埋め尽くされていますがT 細胞が認識できないだけです。したがって、リンパ球の移動パターンは、適応免疫系の効率的な活性化を保証するだけでなく、自己抗原に対する寛容性を維持する上でも重要な役割を果たすことは一切ないのです。
寛容誘導二次リンパ器官の話も嘘ですがどのように又何故ウソなのかを一つずつ説明していきましょう。「自己免疫疾患」は存在しないのに「自己免疫疾患」は存在するとウソをついてしまうとウソがウソを新たに生み出し二進も三進もいかなくなってしまう典型的なのです。嘘で塗り固めた理論が「自己免疫疾患」なのです。この嘘が重大な犯罪であるのはお金を儲けるために患者を死ぬまで苦しめるからです。だからこそこれほどまでにしつこくうその根源を追求しているのは自分自身が15歳から悩みはじめ三つ目の大学である京都府立医科大学に入りなおして医学を勉強しなおしたのですが規制の医学には私の病気を治す治療法は一切ありませんでした。絶望が続きましたが中国医学を漢文ですべて解読できた漢方薬局を経営していた素晴らしい人物の娘と出会いがあり23年間苦しんできた慢性頭痛などのヘルペスによる症状がすべて消えてしまったのです。それまではヘルペス脳炎や右目のヘルペス性網膜炎、右目の強度視力障害、ヘルペス性自律神経障害、尋常性乾癬、軽度の多発性硬化症などの様々な症状がありました。漢方生薬を大量に飲み始めてからまず頭痛が減り免疫学の勉強に集中が可能となり何故漢方煎じ薬が私の原因不明といわれていた様々な症状は漢方煎じ薬で消えてしまったのかの答えを出し始めたのです。症状が改善し始めたのは漢方煎じ薬が免疫を上げたとわかりだしたのです。何故ならば京都府立医科大学でも用いられていた治療薬は一回だけのワクチンと一時的に用いる抗生物質以外はすべて免疫を抑制して症状を一時的に除去するだけですべての病気は治っていないことに気が付いていたからです。京都府立医科大学のいわゆる標準医療に絶望し苦しみました。そのような間違った私の気は治らないからです。最悪の治療薬は正にステロイドであることも誰も教えてはくれませんでしたがステロイドを長年使ってきた患者から学びました。自分の長年苦しんできた病気の原因もわからないどころか治せないだけでなく他の難病といわれる様々な患者の病気が治っていないのに絶望して意図的に留年を繰り返しました。生活は塾をやっていたので食つなぐことはできました。自殺未遂も3回繰り返しましたが無理でした。とにかく自殺が無理なので生き延びるためにも医者になったことが漢方薬の出会いがあったのです。京都府立医科大学で学んだ医学は病気を治すのに何の役にも立たないのですが卒業してから漢方医の娘と結婚することで私の人生は蘇生できたのです。他の世界中の医者が治せない自己免疫疾患のすべてを完治させることができました。
胸腺は、骨髄に存在する造血幹細胞から誕生したT細胞が移動し、成熟する一次免疫組織です。多くの造血幹細胞からつくられて成熟していきますが、T細胞だけは造血幹細胞で作られたままの状況で、胸腺へと移るという特徴があります。 つまり、胸腺はT細胞を作る上でなくてはならないものなのです。ナイーブT細胞とヘルパーT細胞の違いは何ですか?
ナイーブT細胞はこれまで抗原に出会ったことのないT細胞で、MHCクラスⅡと抗原ペプチド複合体による抗原提示に対応するものはナイーブヘルパーT細胞です。 T細胞は骨髄で未熟型が作られ、胸腺で成熟してナイーブT細胞となって全身のリンパ節を巡回しています。T細胞は、表面にT細胞レセプター(T cell receptor, TCR)とよばれる抗原受容体を発現する細胞です。TCRは、B細胞の抗原レセプターである免疫グロブリンと同様、V(D)J組換えにより抗原認識多様性を生み出します。一方、TCRは免疫グロブリンと異なり、抗原分子そのままに結合するのではなく、抗原提示細胞上のMHC分子に提示されたペプチド抗原に結合しなければなりません。
自己の成分を敵とみなさないので自己免疫疾患を絶対に起こさせない樹状細胞とは何か?
樹状細胞(dendritic cell略してDC)は、病原体を見つけ出しそれに続く適応免疫応答の活性化において主役を果たします。DCは、主要組織適合遺伝子複合体(MHC)分子に乗せた病原体の抗原ペプチドを提示することにより、適応応答を開始してT細胞を活性化して分化を誘導します。また、DCは免疫応答を増強および調節するサイトカインと増殖因子を分泌します。ナイーブT細胞の活性化におけるそれらの役割に加えて、DCは制御性T細胞の分化ならびにT細胞が自己免疫疾患を起こさないようにしています。敵を見分ける重要な歩哨細胞として、それらは全身、特にリンパ器官ならびに腸や皮膚などに常在します。
樹状細胞は自然免疫と獲得免疫の橋渡しをします。DCはそれらの環境を継続的にサンプリングし、炎症シグナルがあると活性化します。DCの活性化や成熟は、病原体関連分子パターン(PAMP)または細胞が病原体によって傷つけられたときに生じるダメージ関連分子パターン(DAMP)をTLR (Toll様受容体)などのパターン認識受容体(PRR)により認識すると活性が起こります。DCは、表面およびエンドソームのToll様受容体(TLR)、C型レクチン、および細胞質ゾルセンサーなど多岐にわたるPRRを発現します。エンドソームとはエンドソーム (endosome) はピノサイトーシス(飲作用)によって形成された一重の生体膜からなる小胞で、 ほぼ全ての真核細胞が持ち、細胞外の分子の取り込みや、細胞表面の分子のソーティング(えり分け)に関わります。 その結果エンドソームに取り込まれた一部の分子は再利用され、小胞輸送によって細胞膜へと輸送される。PRR(パターン認識受容体)が活性化されると、DCは代謝、細胞、および遺伝子転写の変化を起こし、Tリンパ球の効率的な活性化因子に成熟するのがDCなのです。成熟したDCはMHCクラスII、共刺激分子、および炎症誘発性サイトカインなど抗原提示装置の発現を増加させ(アップレギュレーション)、二次リンパ組織のT細胞領域に遊走し、そこで抗原特異的T細胞をプライム(抗原刺激)して活性化するのです。成熟した樹状細胞は、形態学的、代謝的、および機能的な変化を起こすので、リンパ組織に遊走して抗原特異的T細胞応答を開始することができます。
胸腺は今なお不明な点が多いので復習しておきます。胸腺の構築は,そこに分布する血管系の骨組みのうえに胸腺上皮細胞の網目とマクロファージや樹状細胞(dendriticcellsDC)などのストローマ細胞(stromalcel1)が存在することによってできています。この胸腺のなかで骨髄からやってきた胸腺前駆細胞が増殖し,成熟し末梢に動員されてT細胞として機能します。そのT細胞は自己の主要組織適合遺伝子複合体(MHC)分子に乗せられたいろいろな非自己のペプチド抗原などに反応できるレパートリーをもち,かつ自己の成分には反応することはないようなT細胞抗原レセプター(T cell receptor,TCR)を発現するように胸腺のなかで教育されるだけで自自己の成分に対する自己寛容の教育はしません。また,胸腺で作られた胸腺細胞のうち95%近くは胸腺内で死滅し,有用なT細胞として末梢に動員される細胞はわずか数%であるのは価値のあるT細胞抗原レセプター(T cell receptor,TCR)を持たせることが極めて困難であるので生き残る正常な病原体だけを敵として認識できるTCRをもったT細が少ないのです。因みに骨髄で正常な機能を持ったB細胞抗原レセプター(B cell receptor,BCR)を持ったB細胞を作られても生き残るのは数%しかないのと同じなのです。
T細胞が胸腺のなかで形成される仕組みは現在でも不明な点が多いのです。だからこそ今なお、この世にありえない自己免疫疾患がない教育が胸腺で行われているとウソが続いているのです。胸腺については不明といわれている点が4つあります。(1) 胸腺においては造血系の幹細胞からTリンパ球のもとになる胸腺細胞しか作られないのは,すでに胸腺にやってくる幹細胞がそのように運命づけられているのか? (2)作られるT細胞はあらゆる抗原に反応できるレパートリーをもった多様なT細胞抗原レセプター(TCR)を発現するT細胞を作り全身に送り出します。て自分の成分にまで反応するものは送り出していない仕組み(negative selection)があるとされているがそれはほんとうにそうなのかとかは100%ウソです。(3)20才を過ぎると胸腺は退縮して脂肪組織になるのですが,胸腺はもうその頃以降にはほんとうにあまり役立っていないのだろうかとかという疑問に対する答えは後述します。簡単な答えはメモリーT細胞があるので必要がないのです。(4)作られた胸腺細胞が末梢に出かけていってT細胞になるわけだが,どのようなルートで、胸腺の静脈を経由するのか輸出リンパ管を経由して出ていくのかの疑問で4つの疑問です。答えは胸腺の静脈も輸出リンパ管の二つの出口があるのでどちらも利用しています。
胸腺で生まれたての抗原に出会ったことがないナイーブ T 細胞の制限された交通パターンは、特に存在しません。ナイーブ T 細胞の仕事は如何に早く抗原提示細胞である樹枝状細胞に自分に合う抗原をいち早く出会うかが仕事の一つです。何故ならばリンパ管のリンパ液は心臓に戻って血液と合流するからです。
二次リンパ器官とリンパ管とは何かを詳しく説明しましょう。
特定の病原体である侵入者に対して特異的な TCR を持つ T 細胞は実は 100 ~ 1,000 個しかなく、これらの T 細胞が活性化されるには、その侵入者を「見た」抗原提示細胞である樹枝状細胞と接触する必要があります。これらの特定の敵だけを認識できる T 細胞と 死の敵を捕まえてくれるAPC は体中に広がって存在しているので、病原体の侵入が完全に制御不能になる前に100 ~ 1,000 個の中の特定のT 細胞とAPCとが出会い起こる可能性は非常に低いのです。この出会いを妥当な確率で機能させるために、免疫系には「T 細胞とAPCとの会合場所」である二次リンパ器官が存在しています。最もよく知られている二次リンパ器官はリンパ節なのです。
家庭には 2 つの配管システムがあります。1 つ目は蛇口から出る水を供給します。これは加圧システムで、圧力はポンプによって供給されています。二つ目はシンク、シャワー、トイレの排水口を含む別の配管システムがあります。この 二つ目のシステムは加圧されていません。水は排水口を流れて下水に流れます。これらの家庭の2 つのシステムは、最終的に廃水がリサイクルされて再利用されるという意味でつながっている2 つの配管システムがあります。
人間の血管とリンパ管の二つの配管はこれによく似ています。一つは心血管系の心臓のポンプによる加圧システム があり、血液はこの心臓のポンプによって体中に送り出されます。しかし、私たちには別の配管システムもあります。リンパ系です。このリンパ系のシステムは加圧されておらず、血管から組織に漏れたリンパ液という体液を毛細リンパ管に吸収して組織からリンパ管に組織からを排出してしまいます。このシステムがなければ、組織は体液であるリンパ液で満たされ、むくみや腫れがひどくなり体全体がむくんでしまい、むくみが増えるばかりとなります。
リンパは上半身ではなく主に下半身の組織から下半身のリンパ管に集められ、筋肉の収縮により、一連の逆流を防ぐために一方向弁を通って上半身へと下半身のリンパ管によって輸送されます。このリンパ(リンパ液)と体の胸より中央寄りの上の上半身の左側からのリンパは胸管に集められ、左鎖骨下静脈の血管に排出されて心臓に戻し、そのリンパは血液と混ざって心臓のポンプ作用によって血管で再び全身に循環されます。同様に、上半身の右側からのリンパは右リンパ管に集められ、右鎖骨下静脈に排出され心臓に戻ります。下の「リンパ管とリンパ節の分布」の図から、組織に出たリンパが血管に再吸収されて血液と再び心臓で合流するために曲がりくねって戻るときに、リンパ管の一連の中継地点である黒い点印のリンパ節を通過することお分かりになるでしょう。
右の図の英語の日本語の訳
Lymph Node リンパ節
Right Lymphatic Duct 右リンパ管
Left Subclavian Vein 左鎖骨下静脈
Thoracic Duct 胸管
Lymph Node リンパ節
Lymphatic Vessel リンパ管
Lymph Node リンパ節
右の絵は「リンパ管とリンパ節の分布」
人間の体には、非常に小さいものから芽キャベツほどの大きさのものまで、約 700個のリンパ節があります。ほとんどのリンパ節は、リンパ管でつながれた「鎖」状に並んでいます。細菌やウイルスなどの侵入者はリンパによって近くのリンパ節に運ばれ、組織内で外来抗原を拾った抗原提示細胞はリンパ節に移動してそのウイルスの積荷を提示します。一方、B 細胞と T 細胞はリンパ節からリンパ節へと循環し、「運命づけられた」自分にピッタリ合う抗原を探します。つまり、リンパ節は実際には「デート バー」として機能し、T 細胞、B 細胞、APC、抗原の四つがすべて集まってコミュニケーションと活性化を行う場所です。細胞と細胞のコミュニケーションとは何でしょうか?多細胞生物である人体の社会秩序を保つためには細胞同士のコミュニケーションが大切です。このコミュニケーションにいつも使われるのはシグナル分子といわれるコミュニケーション専用の化学物質がよく使われます。このシグナル分子を別の細胞が受容体で受け取れば細胞間のコミュニケーションが成立します。
これらの細胞と抗原をリンパ節の小さな容積内に集めると、それらが相互作用して適応免疫系を効率的に活性化する可能性を大幅に高めるためにリンパ節は存在しているのです。リンパ節はあくまでも未熟なT 細胞、未熟なB 細胞、APC、抗原の四つがすべて集まって四つの免疫系が集まってコミュニケーションを行って未熟なT 細胞、未熟なB 細胞の活性化を行う場所です。決して病原体や癌細胞を殺戮するためにあるのではないのです。二次リンパ組織であるリンパ節はいわば教育機関であって戦場ではないのです。それでは何故がんのリンパ節転移が頻繁に起きるのでしょうか?答えは極めて簡単です。強力な兵器や勇猛な戦士がいないのでリンパ節に隠れるためなのです。詳しい説明は後述します。乞うご期待!!!リンパ節の存在価値は初心なT細胞やB細胞を一から教えて大人にする教育機関なのです。教育が終わった後に病原体のいる社会組織に出て戦いを準備する学校に過ぎないのです。だから癌細胞やヘルペスにとっては学校である骨髄やリンパ器官ほど安全な場所はないのです。
リンパ液が流れる管のことをリンパ管と呼びます。組織に出たリンパ液を元のリンパ管に戻し最後は心臓に戻すのです。心臓というポンプから押し出された血液とリンパ液は心臓のポンプ作用で組織に流れ出ていきます。組織に出たリンパ液をそのままにしておくと、リンパ液の量が組織で増えすぎてしまい、むくみや腫れの原因となる可能性があるため、リンパ管が組織に出たリンパ液を血管に戻すことで、正常なバランスを保っているのです。また、血管の血液が体内を円のように循環するのに対し、リンパ液は心臓から血液と一緒に出ていき血液と組織まで運ばれて組織にでていきます。毛細リンパ管の内皮細胞同士がボタン様結合で繋がって特徴的な柏の葉状の輪郭を示し,ボタン同士の間の広く開いた細胞間隙から免疫細胞・脂質・異物・癌細胞などの大型物質を取り込むことができます。リンパ管は末梢組織内で血管と吻合しない網状ネットワークとなっていて,固有の役割を果たすことができる.
リンパ管は体の末端の組織で途切れており、組織に出たリンパ液は自然にリンパ管の内皮細胞間隙から吸収されますが血液の様に循環もすることもできないので心臓で血液と混ざり血液に体内を組織に出るまで循環させてもらうのです。組織にいるウイルスや細菌などの病原体はリンパ管に吸収されて700個以上にも及ぶリンパ節を通過して心臓に戻るのです。心臓に戻る時にリンパ液が逆流しないように弁が付いており、リンパ液は心臓に向かうだけの単一方向にしか流れない点も特徴的です。
組織で病原体を抗原提示細胞である樹枝状細胞を捕まえたこの病原体をB細胞やT細胞に
提示して合うB細胞やT細胞を探すために700個以上にも及ぶリンパ節を旅しまわるのです。するのです。つまりリンパ節とはB細胞やT細胞と抗原提示細胞である樹枝状細胞の三つが出会って樹枝状細胞がB細胞やT細胞を活性化するためにデート(密会)するためのバーなのです。つまり直径3センチ位の芽キャベツや豆粒の大豆のような小さいリンパ節が人体の全身に700個もあるのはB細胞やT細胞と抗原提示細胞である樹枝状細胞の三つが集まる目的は自然免疫の樹枝状細胞が適応免疫のB細胞やT細胞と相互作用してB細胞やT細胞を活性化する確率を高めるための教育機関というべきなのです。何故がん細胞はリンパ節に転移したがるのでしょうか?がん細胞はリンパ節で免疫機構により攻撃を受けるといわれていますが本当でしょうか?そもそも癌細胞は人間の遺伝子23500個の中の2種類の癌関連遺伝子は合計で800個近くあります。この2種類の癌関連遺伝子のワンセットの二つが変異起こしてこの2個の異常なタンパク質ができた細胞を抗原提示細胞である樹枝状細胞のTLRが危険なシグナルと認識して敵が来ましたとB細胞やT細胞に伝えて2個の異常なタンパク質を攻撃して炎症が生まれると思いますか?無理です。遺伝的多型とみなし同じ種の中でみられる遺伝的な個性として広くみられる遺伝子のばらつきで作られる蛋白質は異物としてみなすことをしないので病気を起こすことにはならないのです。顔の違いや性格の違いは遺伝子的多型のばらつきで作られる蛋白の違いですから病気ではないのです。美人でないとか頭がよくないというのは病気ではないのです。それでは癌の下人は何ですかに対する答えはヘルペスであるのです癌細胞には非常に多くのヘルペスが増殖しているのです。このヘルペスは病原体のウイルスですから免疫の力が弱いリンパ節にこぞって逃げ隠れするために転移しやすい癌の原発巣の最も近いしかも免疫の弱いしかもヘルペスの怖さを何も知らないナイーブT細胞のいる所属リンパ節に転移し始めるのです。癌についてはロイアルレイモンド博士のここを読んでください。
何故がん細胞はリンパ節に転移したがるのでしょうか?癌では初期からリンパ節転移が観察されるのは何故でしょうか?リンパ組織に侵入する機序は詳しくわかっていないようですので私が説明してあげましょう。免疫のイロハがわかっていないナイーブTリンパ球ばかりが活性化のためにひしめいているリンパ節は全身で700ヶ所あり、そこで癌細胞が転移してもナイーブTリンパ球は癌を殺す力はないので癌細胞は生き残り続けます。リンパ液の流れも圧が掛からないために緩やかでさらにリンパ節の周りには筋肉の収縮もないのでリンパの流れが更に弱くなるので居座ることになり癌細胞の増殖に伴って増殖に専念できます。がん細胞もヘルペスがリンパ節で増殖し固定的な転移巣となりリンパ節の辺縁洞や髄洞にヘルペス性癌を貪食しようとして血管から集まった大食細胞すぎてリンパ節が腫れてくるのです。さらに増えすぎた癌細胞は輸出リンパ管から次のリンパ節まで転移したり、静脈を通じて他の組織に転移することもあるのです。転移した先で増殖し、やがて正常細胞よりもがん細胞が勢いをつけ栄養を得るようになってしまうことで、それぞれの臓器が正常な働きをおこなうことができなくなることもあります。
リンパ節には様々な臓器からの癌転移が見られます。 肺癌の場合、血液やリンパ液の流れに乗って癌が広がるため、リンパの流れが密集しているリンパ節に転移することが多いそうです。 乳癌ではリンパ節に転移する確率が最も高いと言われています。 胃癌の場合は、癌細胞が胃の粘膜を侵食していくことでリンパ節に転移します。
がんは大きく成長すると、一部のがん細胞がリンパ液や血液に乗って他の臓器に転移します。
細胞はなぜ死なないのでしょうか?
がん細胞ではたいてい、不死化をもたらすテロメラーゼ(telomerase)と呼ばれるテロメア合成酵素が活性化しており、この酵素の働きによってテロメアが安定に維持されます。がん細胞が無限に分裂出来るのはこのためです。
テロメラーゼ(telomerase)とは何でしょうか?
左図はテロメラーゼによるテロメア配列付加の模式図です。ヒトのテロメラーゼは染色体末端DNAの 3’側に6塩基配列 TTAGGGを付加する。左下図は付加された配列をテンプレート(鋳型)としてDNAポリメラーゼが相補鎖を合成する。下左図は末端複製問題とテロメアの図です。DNAはDNAポリメラーゼ(青丸)によって複製されるが、最末端のプライマー(赤線)部分は複製されない。このため、複製のたびにDNAは短縮する。これが「末端複製問題」である。下右図は生殖細胞やがん細胞ではテロメラーゼによって末端部分の複製が行われる図です。テロメラーゼ活性がない体細胞では分裂ごとに短縮がおこり、一定以上短くなると分裂を停止し細胞老化が起こる。
テロメラーゼ (英: telomerase) は、真核生物の染色体末端(テロメア)の特異的反復配列を伸長させる酵素。テロメア伸長のテンプレート(鋳型)となるRNA構成要素と逆転写酵素活性を持つ触媒サブユニットおよびその他の制御サブユニットによって構成されている。
テロメラーゼ活性が低い細胞は、一般に細胞分裂ごとにテロメアの短縮が進み、やがてヘイフリック限界と呼ばれる細胞分裂の停止が起きる。テロメラーゼは、ヒトでは生殖細胞・幹細胞・がん細胞などでの活性が認められ、それらの細胞が分裂を継続できる性質に関与している。このことから、活性を抑制することによるがん治療、および活性を高めることによる細胞分裂寿命の延長、その両面から注目を浴びている。
酵素によりテロメアが伸長されることは、1973年にアレクセイ・オロヴニコフ (Alexey Olovnikov) によって最初に予測された。彼はまた細胞老化に関するテロメア仮説およびがんとテロメアの関連について示唆を行った。
1985年にカリフォルニア大学のキャロル・W・グライダーとエリザベス・H・ブラックバーンは、テトラヒメナからこの酵素を単離したことを公表した。グライダーとブラックバーンはジャック・W・ショスタクと共に、テロメアとテロメラーゼに関する一連の研究で、2009年ノーベル生理学・医学賞を受賞した。
概要
構造と機能
テロメラーゼはテロメア配列の鋳型となるRNAと逆転写酵素、その他の制御サブユニットからなる複合体である。RNA構成要素はTERC (Telomere RNA Component, TRとも表記) 、逆転写酵素はTERT (Telomere Reverse Transcriptase) と呼ばれる。このRNAの長さはテトラヒメナで159塩基長、ヒトで451塩基長、出芽酵母で約1,300塩基長と様々である。逆転写酵素の活性部位はRNA型トランスポゾンがコードするそれと相同性がある。過剰発現の実験から、テロメラーゼ活性自体はRNAと逆転写酵素の二つの構成因子で十分であることがわかっているが、テロメラーゼは生体内において巨大な複合体 (1MDa以上) を形成しており、正常な機能には他の構成サブユニットも必要である。
ヒトのテロメラーゼは、TERT、TERC、ジスケリン (dyskerin) 、TEP1などのサブユニットによって構成されており、それらは異なる染色体上の遺伝子座にコードされている。TERT翻訳産物(タンパク質)は、非翻訳RNAであるTERCと一緒に折りたたまれる。TERTは一本鎖テロメア反復配列を付加できるように染色体の周囲を覆う二股の構造をとる。TERTとテロメアの鋳型を含むTERCは隣接している。ヒトTERCでは鋳型配列領域は 3′-CAAUCCCAAUC-5’であり、これを元にTERTはテロメアの3’側へ塩基を付加する(脊椎動物では6塩基配列5′-TTAGGG-3′(GGTTAG)を付加するが、他の生物では別の配列)。テロメラーゼは、この塩基付加を繰り返し、染色体のテロメアの伸長を行う。
コクヌストモドキ (Tribolium castaneum) TERTのタンパク質構造の詳細な解析が、2008年に行われた。このTERTは4つの保存されたドメイン(TRBD, fingers, palm, thumb)を含むタンパク質であり、レトロウイルスの逆転写酵素・ウイルスのRNAポリメラーゼ・バクテリオファージのDNAポリメラーゼ(ファミリーB)と共通の特徴を持つ環状構造をとっている。
テロメアおよびテロメラーゼの分子機構に関する実験には均一な細胞群を用いることが求められるため、主に出芽酵母やテトラヒメナといった単細胞生物、および哺乳類では培養細胞を用いて研究が行われている。テロメラーゼは細胞周期のS期(DNA合成期)にテロメアに誘導されて機能する。出芽酵母の研究では、テロメラーゼは細胞内で最も短いテロメアから優先的に伸長させていくことがわかりつつあり、長すぎるテロメアには抑制的に働く機構が見いだされている。
活性
テロメラーゼの活性については、生物・組織・細胞の種類によって異なることが知られている。真核単細胞生物は例外なくテロメラーゼ活性を持ち、真核多細胞生物では生殖細胞にはテロメラーゼ活性があるが体細胞での活性はさまざまである。植物においては調べられた殆どの体細胞でテロメラーゼ活性があり、このことが株分けなど栄養生殖でほぼ無限に増殖できる不死性を持つ一因になっていると考えられている。ヒトでは生殖細胞・幹細胞以外での活性がほとんど見られないが、同じ脊椎動物でも魚類・マウス・チンパンジーでは体細胞でのテロメラーゼ活性が観察されている。
ヒトでのテロメラーゼ構成要素の発現をみると、RNA構成要素TERCは体細胞でも発現しており、酵素活性は触媒サブユニットTERTの発現で調節されている。ヒト培養細胞でゲノム中のTERTを強制発現をさせることは困難であるが、人為的に別のプロモーターを付加したTERTを導入することにより細胞の不死化を行うことができる。ヒトのがん組織の多くではテロメラーゼが大量に存在しており、がん細胞の不死化の原因の一つと考えられている(一部のがん組織はテロメラーゼ陰性)。また、生殖細胞は個体を超えて世代を継続させる一種の不死性を持つが、テロメラーゼが恒常的に発現していることがその一因となっている。
臨床関連
がん
ヒトのがん組織の多くではテロメラーゼの活性化がおきており、その観察される割合は肺がんの80%から食道がんの95%に及ぶ。一方、テロメラーゼ活性の制限要因であるTERTの発現が見られない腫瘍も観察されており、それらではALT(Alternative Lengthening of Telomeres)と呼ばれるテロメア長を維持する別の機構が見出されている。
ヒトのがん細胞とテロメラーゼとの関係についての説明は以下のようなものである。
- ヘイフリック限界に達した正常細胞は、p53遺伝子やRb遺伝子などのがん抑制遺伝子の働きによって細胞分裂停止が起きる。
- がん抑制遺伝子に突然変異が起きた細胞は、上記の限界を超えて分裂を続け、テロメア短縮もさらに継続する。
- 通常はこれらの異常細胞は、p53経路による細胞死か、染色体の構造異常による細胞死を迎える。この過程の中で、ゲノムの不安定化が起こり、さらにさまざまな変異を誘発することがある。
- 上記の中の一部には、TERTの恒常的発現を獲得する変異細胞があり、それらはテロメラーゼの活性化を通して染色体を安定化させ、腫瘍形成に至る。
このようにテロメラーゼ遺伝子自体ががん化の直接原因ではないが、がん組織の形成増殖にとってテロメラーゼ活性化が必要である場合が多いため、テロメラーゼを標的とした抗がん剤の開発が行われている。その例として、ヒトTERC(hTR)を標的とするGRN163/GRN163L(ジェロン社)、テロメア短縮を誘導するBIBR1532(ベーリンガーインゲルハイム社)、テロメア構造の安定化に作用するテロメスタチンなどがある。
個体老化
遺伝的な要因による早老症として、ヘリカーゼ遺伝子を原因とするウェルナー症候群・ブルーム症候群 (Bloom syndrome) 、DNA修復に関連するキナーゼ遺伝子ATMを原因とする毛細血管拡張性運動失調症、ゲノム損傷修復の関連遺伝子を原因とするナイミーヘン症候群 (Nijmegen breakage syndrome) などが知られている。これら早老症患者の細胞では分裂寿命が短くなりテロメア短縮も早まる事例があるとする報告や、さまざまな早老症がテロメア短縮と関連しているとする報告がある。これら遺伝子群のDNA修復機能とテロメア長の維持との正確な関連は判明しておらず、個体老化(早老症)とテロメア短縮との関連について研究が進められている。
テロメラーゼを利用した療法が、ヒトの老化を回避して寿命を延長させることにも使われると考える医師もいる。しかしながら、テロメラーゼの活性化には、細胞老化防止の可能性と、正常細胞のがん化の一因となり個体寿命の短縮化をもたらす可能性があることの両面が指摘されており、アンチエイジングへの応用についての評価は定まっていない。
他のヒトの疾患
テロメラーゼおよびその触媒サブユニットTERTは、がん・早老症以外の疾患との関係も調べられている。
TERTと関係すると考えられている疾患としては、再生不良性貧血、猫鳴き症候群 (Cri du chat Syndrome,CdCS) がある。CdCSは、5番染色体短腕(5p)の末端部分の欠損を含んでいる複雑な障害である。TERTは5p領域(染色体上の位置 5p15)にあり、TERTの欠損はCdCSの原因または関与要因として示唆されてきた。
先天性角化異常症 (dyskeratosis congenita, DCまたはDKC) は、テロメラーゼ活性と関連する骨髄の疾患である。症例の35%はX染色体連鎖(伴性遺伝)の劣性遺伝子疾患であり、5%は常染色体性の優性遺伝子疾患、残り60%は原因不明である。X連鎖劣性の場合はジスケリン(染色体上の位置 Xq28)、常染色体性優性の場合はTERC(染色体上の位置 3q26.2)またはTERTの変異が原因となっている。DC患者は、いろいろな他の徴候だけでなく、異常な網状皮膚色素沈着、白板症(口腔粘膜の白い肥厚)と爪の発育異常として現れるひどい骨髄不全を示す。X連鎖または常染色体性どちらのDC患者でも、同じ年齢のほかの人よりも短いテロメアと欠陥のあるテロメラーゼ活性を示す。常染色体優性DCの患者家系の一つでは、世代が進むにつれ、テロメア短縮の率の増加および発症する年齢の低下現象(表現促進)、すなわち各々の世代での悪化が見られる例も存在する。
転移した癌はなぜ治らないのですか?
転移した癌の治療は、初期の癌に比べて難易度が高くなります。これは、転移した癌細胞が体内の複数の部位に分散しているため、全ての癌細胞を除去することが難しいからです。血液やリンパの流れでがんが転移し全身に行き渡ってしまうと、手術でがんを取り除くことは不可能です。もし仮に手術で取り除こうとすると、手術時間が長くなるだけではなく、取り除く量も多くなり、体への負担が大きく、体力、免疫力を奪ってしまい、逆にがんの勢いが増してしまう可能性がありますといわれていますが癌の原因がヘルペスでありかつ転移も上で説明したようにヘルペスが大きくヘルペスが関わっていることを考慮していないのでロイアルレイモンド博士の「癌光療法」で癌ウイルスであるヘルペスウイルスを殺せば転移癌であろうがなかろうが、いかなる臓器のどんな癌であろうが完治できます。
ガン細胞は体温が何度で死滅しますか?
温熱療法とは 腫瘍を電磁波で体外から癌細胞を標的にして42.5度以上加温する治療です。 人間のどんな細胞であろうが42.5度以上に温度があがると死滅します。
癌細胞は何年で癌になる?
1cmの大きさに成長するまでに要する時間は10年から15年といわれています。1つのがん細胞が1センチの大きさになるまで10年から15年かかりますと言われていますが間違いです。しかし、1センチのがんが2センチになるには、1年から2年しかかかりません。
この説明もヘルペスの増殖と癌の増殖との関連は一切考慮されていませんのが問題になり正確さに欠けます。癌の増え方にヘルペスが絡むと何故正確な癌の増殖の数値が出なくなるのでしょうか?それはヘルペスの増殖は癌患者さんの免疫の強さに大きく左右されるからなのです。勿論がん治療の違いによって癌患者の免疫が以外によって大きく影響を受けるので患者の免疫を落とせば確実にヘルペスが増えてしまうので癌細胞の転移も増えかつ癌関連遺伝子の癌化も増えるので癌の増殖と癌転移の拡大も比例してしまうからですが現代の癌学者やがん専門医は誰も気が付いていないので癌に関する数値は全く信用できないのです。いずれにしろ現代の医療のすべては多かれ少なかれ免疫を落とすことになるので「癌は放置するのが一番長生きできる」という故近藤誠先生の意見は正しいのですが彼は癌の原因はヘルペスであるとかヘルペスが免疫低下で増えるので癌も大きくなるなどは一切知りませんので彼は癌を医療で治すことができなかったのです。ロイアルレイモンド博士や松本博士の様に癌ウイルス(ヘルペスウイルス)が癌の原因であると知っている人は世界中に何人いるでしょうか?誰もいないでしょうね。残念です。現在は医者から完全に見放されて緩和医療しかないと診断され見放されている末期がん患者はともかく良性腫瘍や癌ではあるが転移していない悪性腫瘍の患者は治せるという確信をも持ち始めています。だって38年臨床経験で10人以上の癌患者さんが治ったのは抗ヘルペス剤と免疫を上げる大量の漢方洗剤で完治させたからです。癌を治したすべての患者さんにはあなた自身が完治させたのです。癌になったのはあなたがストレスでステロイドホルモンを出しすぎて免疫を抑えすぎたためにヘルペスを増やしたために二つの癌を作る関連遺伝子を癌化させたために癌になったのです。癌を大きくさせたり転移させたくなかったらストレスを減らしなさい。そうすれば癌の原因であるヘルペスが増えないので癌は治る可能性が生まれるよ。私は細胞の中でヘルペスが増えないようにするための抗ヘルペス剤と増えたヘルペスが細胞から出た時に免疫を上げる漢方薬を出すからね。癌を作らないためにも癌が大きくならないためにも更に転移しないためにも免疫が一番大事だからね。といっただけで癌も自分の免疫でしか治せないのです。現在のすべての病気を治すのは自分の免疫しかないのです。今も昔も免疫を最高度に落とすステロイドホルモンがすべての病気の原因であるヘルペスを増やしてしまうのであるので免疫を落として症状をとる対症療法は病気つくりの最高の薬です。
後天性の遺伝子病である癌も先天性の遺伝子病も後天性の遺伝子病も細胞に感染したヘルペスが核のゲノムの染色体に自分のゲノムを組み込みかつ部分特異的組み換えを意図なくたまたま偶然ではあるけれども遺伝子を組み替えたための必然の結果細胞のゲノムのDNAの配列が変わって異常なたんぱく質を作らざるを得なかったので生じた突然変異によって生まれた遺伝子病です。
リンパ液とは何か?リンパ液とは、血管から出たが体内の細胞に栄養や酸素を届けた後、血管に戻れなかった組織液(水分)の一部です。主成分は、血液の液体物質である血漿とリンパ球ですが、ほかにも傷ついた細胞や、細菌やウイルスなどの異物も一緒に含まれていることがあります。リンパ液は心臓から出口にある静脈に向かって一方向に流れています。最近まで、自己免疫を防ぐ胸腺の唯一の役割は、自己反応性の可能性のある T 細胞を排除することであると考えられていましたが間違いです。しかし、現在では、自己免疫疾患から私たちを守るのに役立つ胸腺の別の機能、つまり自然制御性 T 細胞 (nTreg) の生成があることが明らかになっています。
自然制御性 T 細胞 (nTreg)とは何でしょうか?T細胞サブセットの一つである制御性T細胞(Treg)は自然発生的Treg(naturally occurring regulatory T cell略してnTreg)と誘導性Treg(inducible regulatory T cell略してiTreg)に大きく2つに分類される.自然発生的(内在性)Tregは胸腺内で自然発生し,一方,誘導性Treg(iTreg)はTGF-βの存在下における抗原刺激により末梢血中のナイーブT細胞から分化誘導される.
TGF-βとは何でしょうか?TGF-β〈transforming growth factorβ、訳して、形質転換増殖因子〉は線維芽細胞に対する形質転換活性をもつ増殖因子としてのサイトカインの一種であるが、種々の細胞に対する増殖抑制、分化、遊走、接着、炎症、免疫など、さまざまな作用の発現に関与する。TGF-βはポリペプチドがS-S結合した25kDaの2量体分子であり、TGF-β1~5の5種類のアイソフォームがある。
TGF-βの作用は、細胞によって大きく異なり、上皮細胞、血管内皮細胞、リンパ球などの増殖は抑制するが、線維芽細胞、平滑筋細胞などの増殖に対しては促進的に作用する。卵巣、精巣、副腎などのステロイド産生細胞や筋細胞の増殖には影響しない。また、細胞の分化・遊走に対する作用もあり、上皮系細胞や軟骨細胞の分化促進、骨芽細胞や筋芽細胞の分化抑制のほか、線維芽細胞や炎症性単核球の遊走促進、内皮細胞の遊走抑制などを示す。さらに、コラーゲン、フィブロネクチンなどの細胞外マトリックスの産生を促進し、これらマトリックスを分解するプロテアーゼの産生を抑制する。そのほかに増殖因子、サイトカインに対する作用も認められ、線維芽細胞や平滑筋細胞からのPDGF(platelet-derived growth factor、血小板由来増殖因子)の産生促進や、単球からのTNF-α(tumor necrosis factor-α、腫瘍壊死因子-α)やIL-1の誘導を示すこと、アポトーシスの促進や免疫能の抑制などさまざまな作用がある。
TGF-βは肺線維症や肝硬変などの線維化を伴う種々の病態に関与し、また癌細胞の周辺の細胞においてその産生が亢進していることから、血管新生や細胞外マトリックスの増加や免疫能の抑制を介して癌の進展に寄与している。血管新生もヘルペスの増殖に必要であるのでこの血管新生や細胞外マトリックスの増加といった作用は、ヘルペスとの共存のために血管新生や細胞外マトリックスの増加や免疫能の抑制を介して癌の進展に寄与しているのはヘルペスを殺さないようにしているとも考えられます。ちょうどPD‐1とPDL‐2との関係と同じです。PD‐1とPDL‐2についてはここを読んでください。TGF-βの様に細胞の違いによって促進性と抑制性の相反する機能を有しているサイトカインはヘルペスウイルスと共存するために
免疫細胞に対しては抑制性の働きを示す。このような促進性と抑制性の相反する機能を有しているサイトカインの他の例としてはBcl-2がありヘルペスウイルスと共存するために
免疫細胞に対してはBcl-2は抑制性の働きを示す傾向があるのです。
TGF-βは単球からのTNF-α(tumor necrosis factor-α、腫瘍壊死因子-α)やIL-1の誘導を示すこと、アポトーシスの促進や免疫能の抑制などさまざまな作用がある。
TGF-αは、ヒトではTGFA遺伝子にコードされるタンパク質で、TGF-αはEGF(上皮成長因子)ファミリーのメンバーであり、分裂促進因子としての活性を有する。このタンパク質は上皮成長因子受容体(EGFR)のリガンドとなる成長因子であり、TGF-αの結合によって活性化されたEGFRは細胞増殖、分化、発生のためのシグナル伝達経路を活性化する。
メタロプロテイナーゼとはMetalloproteinaseは、触媒機構に金属が関与するプロテアーゼである。プロテアーゼ (protease)は、タンパク質をポリペプチドやアミノ酸への分解を触媒する 加水分解酵素の総称である。ペプチダーゼ (peptidase) やプロテイナーゼ(proteinase)とも呼ばれる。それらは、水が反応して結合を壊す加水分解によってタンパク質内のペプチド結合を切断する。筋形成として知られる、胎児の発達の過程における筋細胞の融合に重要な役割を果たすメルトリンがその一例である。ほとんどの金属プロテアーゼは亜鉛を必要とするが、コバルトを用いるものもある。金属イオンは、3つのリガンドを通してタンパク質に配位する。
腫瘍壊死因子は、固形がんに対して出血性の壊死を誘導する因子として発見されたサイトカインです。TNF とTNFR(TNF Receptor)との結合により、標的細胞のアポトーシス誘導、炎症惹起などの生理作用が発現します。また炎症反応が関わる疾患である関節リウマチ、乾癬、敗血症や骨リモデリングが関わる骨粗鬆症にもTNFファミリーが関与することが報告されています。
TNF-α(tumor necrosis factor-α略してTNF-α、腫瘍壊死因子-α)とは腫瘍壊死因子(TNF-α)は、炎症性サイトカインでメタロプロテイナーゼの作用によりマクロファージをはじめ炎症、発生、分化など広範な生物活性をもつ。
TNFファミリーとは腫瘍壊死因子受容体スーパーファミリーは、細胞外システインリッチドメインを介して腫瘍壊死因子に結合する能力を特徴とするサイトカイン受容体のタンパク質スーパーファミリーです。神経成長因子を除いて、すべてのTNFは典型的なTNF-αと相同です。それらの活性型では、TNF受容体の大部分は原形質膜で三量体複合体を形成します。
IL-1とは重要な炎症誘発性サイトカインでIL-1はIL-1αとIL-1βに細分化されます。IL-1βは強力な炎症誘発性サイトカインであり、微生物の分子に応答して主にリンパ球、マクロファージ、および単球により誘導されます。リンパ球、マクロファージ、および単球がウイルスに感染すると、パターン認識受容体(PPR)とToll様受容体(TLR)が発現し、IL-1βの発現増強につながります。IL-1βはCD4+細胞を刺激し、Th17細胞に分化させます。IL-1ファミリーの刺激効果に加えて、IL-1サイトカインの発現を阻害または抑制できるメンバー(IL-1R、aおよび2)もあります。IL-1Raは、好中球、マクロファージ、単球、および肝細胞から分泌され、炎症の抑制を狙いとしています。しかし、IL-1βの発現を効率的に阻害または抑制するためには、IL-Raの発現を最大1,000倍まで増やす必要があります
IL-6とはIL-6は多面作用性のサイトカインであり、免疫系に影響を与えるだけでなく、他の生体系や、細胞増殖の調節、遺伝子活性化、増殖、生存、および分化などの多くの生理学的事象にも作用します。IL-6は、単球、線維芽細胞、内皮細胞など、さまざまな種類の細胞により産生されます。刺激を受けると、IL-6は、マクロファージ、T細胞、B細胞、肥満細胞、グリア細胞、好酸球、ケラチノサイト、および顆粒球など多くのさらなる種類の細胞により分泌されます。IL-6は、いくつかの種類の白血球、および肝臓での急性期タンパク質の産生を刺激します。特に、B細胞の抗体産生細胞(形質細胞)への分化誘導において重要です。IL-6がその受容体に結合すると、JAK(Janusキナーゼ)のキナーゼの活性化やRasを介したシグナル伝達の活性化などの細胞事象が開始されます。
TGF-βは肺線維症や肝硬変などの線維化を伴う種々の病態に関与し、また癌細胞の周辺の細胞においてその産生が亢進していることから、血管新生や細胞外マトリックスの増加や免疫能の抑制を介して癌の進展に寄与していると考えられている。この血管新生や細胞外マトリックスの増加といった作用は、創傷治癒にも関与している。
胸腺では、CD4+ T 細胞のサブセットが自然制御性 T 細胞になるように選択されます。この選択は胸腺髄質で行われ、mTEC(髄質胸腺上皮細胞) と TDC(thymic dendritic cell胸腺樹枝状細胞)の作用が必要です。現在の考え方では、これらの細胞によって提示される自己抗原に対する受容体の親和性が強い CD4+ T 細胞は排除され、mTEC(髄質胸腺上皮細胞) または TDC(胸腺樹枝状細胞) によって提示される自己抗原に対する受容体の親和性が弱い T 細胞はヘルパー T 細胞として生き残るように選択されますのも間違いです。何故ならば樹枝状細胞は自己成分を提示しないからです。自己である自己抗原などと言う抗原は存在しないからです。また、自己抗原に対する「中程度の」親和性を持つ CD4+ T 細胞は、天然の制御性 T 細胞になるように「誘導」されます。ただし、この文脈で「中程度」が正確に何を意味するのか、またこれらの T 細胞がどのように「誘導」されるのかは不明なのは勝手な空想であるからです。CD4+ T 細胞が nTreg になるように選択されることについてわかっていることの 1 つは、これらの細胞が何らかの方法で Foxp3 と呼ばれる遺伝子を発現するように誘導されることです。この遺伝子は、nTreg 細胞に制御特性を与えるのに役立ちます。胸腺で生成された後、天然 Treg はパスポート (接着分子) を受け取り、リンパ節やその他の二次リンパ器官に入ることができます。実際、循環している CD4+ T 細胞全体の約 5% が制御性 T 細胞です。二次リンパ器官で、天然 Treg が抗原提示細胞によって提示された同族の自己抗原に遭遇すると、活性化される可能性がありますも嘘です。すべて空想の話しです。活性化された nTreg 細胞は、潜在的に自己反応性の T 細胞の活性化を抑制できます。これがどのように達成されるのかは、まだ不明なのも全くあり得ないからです。考えられるメカニズムの 1 つは、Treg 細胞が抗原提示細胞によって提示された同族抗原を認識すると、その APC 上の共刺激分子の発現を減らすように作用することです。これにより、APC が同じ自己抗原を認識する可能性のある自己反応性エフェクター T 細胞を活性化することがより困難になる話しも観念の遊びにすぎません。自己免疫疾患はないからです。
前回の講義では、別の種類の制御性 T 細胞である誘導性制御性 T 細胞について説明しました。誘導性制御性 T 細胞と天然制御性 T 細胞はどちらも Foxp3 タンパク質を発現しますが、抑制活動のターゲットは異なるようです。天然制御性 T 細胞の役割は、自己抗原に反応して自己免疫を引き起こす可能性のある T 細胞から保護することですが、誘導性制御性 T 細胞の主な機能は、免疫系が外来侵入者に対して過剰反応しないようにすることです。この外来侵入者はヘルペスでありヘルペスと過剰反応しても殺し切れないので共存の道を探ろうとしているのが誘導性制御性 T 細胞の主な機能なのです。
天然 Treg(天然の制御性 T 細胞) については未解明の部分がたくさんありますが、自己免疫疾患から私たちを守る上で重要な役割を果たしていることは明らかですと相変わらず根拠も語らず明らかですという医学者の頭がどうなっているのかかちわって見たいものです。実際、Foxp3 タンパク質の機能を損なう変異を持つ人間は、攻撃的な自己免疫疾患に苦しみ、若くして死亡します。この世に存在しない疾患で死ぬことも無いのです。この患者無くなったのも自己免疫疾患は免疫疾患は一生治らない病気だから間違った免疫の働きを抑制するステロイドを大量に使って自己免疫疾患といわれる病気の原因であるヘルペスを無限大増やしてその結果わけのわからないあらゆる症状が出現して治療の使用がなくなり医者が患者を殺してしまっただけのことなのです。
現代の病気の原因は免疫では絶対に殺し切れない従って一度感染してしまってもワクチンの効果に必要な免疫記憶細胞もできないのでワクチンも人工的には効果がなくしかも免疫が高いときは宿主細胞の核の染色体のゲノムの潜伏感染するときに染色体の遺伝子であるDNAを切断してヘルペス自身のゲノムをランダムに組み込みかつ遺伝子の組み換えを自由自在にやってしまい遺伝子を突然変異を起こし癌も起こし原因不明な遺伝子病も作りあらゆる疾患を起こしてしまうのです。しかも患者の免疫が落ちると細胞が二つに分裂するときに結合していた2倍体の23本の染色体が父親と母親の一倍体の染色体の46本になったときにどの染色体に自由に遺伝子を組み込みDNAを部分特異的組み換えによって遺伝子を突然変異させてしまうのです。このように細胞が分裂する時にのみ利用したり新たに作る酵素やタンパク質や機能をヘルペス自身も利用したり奪い取ったりしてヘルペスが持っていない細胞の増殖機構を活用しながらヘルペス自身も何百倍、何千倍にも増えていくのです。このために細胞自身は自分の分裂や2倍の大きさになるために必要な蛋白質や細胞小器官も作れなくなったりするとヘルペスの溶解感染といわれる再活性化とも言われる現象が起こり正常な遺伝子の発現が起こらなくなり細胞変性効果といわれる細胞の変性による形態の変化が生じてしまい様々な細胞機能障害を起こしたり最後は細胞壊死になって死んでしまうのです。細胞変性効果(Cyto‐pathic effect略して CPE)とは何でしょうか?細胞変性効果は、ウイルスの侵入によって引き起こされる宿主細胞の形態変化のことです。ヘルペスウイルス感染により、宿主細胞が溶解する場合や、細胞の複製が阻害され溶解せずに細胞が死滅する場合もあります。これらの現象はどちらもCPEが原因で発生します。ヘルペスウイルスが宿主細胞にこれらの形態学的変化を引き起こす場合、細胞変性(CPE)が起こっているのです。CPEがヘルペスウイルスによって細胞に生じる細胞の形態的な変化には①球状化(rounding)、②収縮(shrinkage)、③屈折性の増加(increased refractility)、④融合(fusion)、⑤凝集(aggregation)、⑥接着性の喪失(loss of adherence)、⑦溶解(lysis)の7つの形態学的な変化があります。これらの目に見える形態的な変化以外に目に見えない変化の代表は様々な変性疾患です。例えばこれらの永代的な変性が様々に組み合わさり、ヘルペスウイルスがんの転移のきっかけとなる癌細胞塊からまず一個の癌細胞が離脱を最初に起こし更に次々と多くの癌細胞が離脱すると癌細胞浸潤をもたらしリンパ所属リンパ節転移となり起こり最後は遠隔転移となってしまうのです。がん浸潤(Invasion)とは
浸潤は、がん細胞が直接伸びてがん内の隣接組織に侵入するプロセスです。これは一般に、循環系またはリンパ系を通ってより離れた場所へがん細胞が広がる転移とは区別されます。しかし、リンパ管浸潤が一般に転移の最初のステップです。 細胞遊走によるがん細胞の浸潤には、主に 2 つのパターンが存在します。
癌についてはここを読んでください。
これらの目に見える形態的な変化以外に目に見えない変化の代表は神経細胞に感染したヘルペスが起こす様々な脳神経細胞変性疾患です。脳神経細胞が変性してしまうと極めて顕著な神経症状が現れ神経変性疾患になります。変性疾患は主に神経変性疾患のことです。神経変性疾患には、パーキンソン病やアルツハイマー病に加え、脊髄小脳変性症や筋萎縮性側索硬化症(ALS、進行性核上性麻痺、大脳皮質基底核変性症、ハンチントン病などのいわゆる神経難病が含まれます。筋萎縮性側索硬化症(筋萎縮性側索硬化症(ALS、ALS、
神経変性疾患とは脳や脊髄にある中枢神経の神経細胞のなかで、ある特定の神経細胞群が徐々に障害を受け脱落し(=神経細胞死)、発症する病気のことです。残念ながらまだ原因はわかっていません。脱落してしまう細胞は病気によって異なっています。
神経変性疾患とは
神経変性疾患とは脳や脊髄にある神経細胞のなかで,ある特定の神経細胞群(例えば認知機能に関係する神経細胞や運動機能に関係する細胞)が徐々に障害を受け脱落してしまう病気です.残念ながらまだ原因はわかっていません。脱落してしまう細胞は病気によって異なっています。大きく分けるとスムーズな運動が出来なくなる病気,体のバランスがとりにくくなる病気,筋力が低下してしまう病気,認知能力が低下してしまう病気などがあります。
スムーズな運動が出来なくなる病気には、脳神経が変性してしまい脳神経の機能が働かなくなり、パーキンソン病,パーキンソン症候群(多系統萎縮症,進行性核上性麻痺など)などがあります。進行性核上性麻痺(progressive supranuclear palsy:PSP)は、中年期以降に発症し、淡蒼球、視床下核、小脳歯状核、赤核、黒質、脳幹被蓋の神経細胞が脱落し、異常リン酸化タウ蛋白が神経細胞内及びグリア細胞内に蓄積する疾患である。アストロサイトの房状アストロサイト(tufted astrocytes)が、PSPに特異的な病理学的所見とされている。神経学的には、易転倒性、核上性注視麻痺、パーキンソニズム、認知症などを特徴とする。発症の原因は不明である。Supranuclearの部位は大脳基底核よりも大脳皮質側に位置する部位に原因のあるという意味です。脳の解剖学に無知な人は何処に大脳基底核があり、大脳皮質がどこにあるかはインターネットで調べたらすぐわかります。脳の話しで「核」と言えば大脳基底核であり「皮質」と言えば大脳皮質のことです
体のバランスが取りにくくなる病気:
脊髄小脳変性症,一部の痙性対麻痺など
筋力が低下してしまう病気:
筋萎縮性側索硬化症など
認知機能が障害されてしまう病気:
アルツハイマー病,レビー小体型認知症,皮質基底核変性症など。皮質基底核変性症は大脳基底核および大脳皮質の神経細胞がヘルペスウイルス感染により脱落し、タウ蛋白という異常なたんぱくが蓄積する変性疾患です。大脳基底核の症状であるパーキンソン病様の運動症状(筋肉の硬さ、運動の遅さ、歩行障害など)と大脳皮質の症状(手が思うように使えない、動作がぎこちないなど)の両者を併せ持つことが特徴です。
神経変性疾患がどのような機序で、なぜ特定の人に起きるのか、始まりはいつなのかも含めてあまりよくわかっていませんが,高齢者に発病しやすい傾向があることから、加齢そのものがリスクであると考えられています.患者さんの家族が同じような症状を持っている事は少ないですが(弧発性),一部の患者さんは血のつながった家族の中に同じ症状、もしくは似た症状を持った方がいて遺伝する事が分かっています(家族性)。最近の研究の進歩により私たちの施設から世界的にも有名なパーキンソン病の原因遺伝子が発見されましたが、さらに多くの遺伝子、蛋白が世界各国で発見されそれらの機能が調べられています.その結果異常な機能を持った蛋白や、必要がなくなった蛋白が分解されずに細胞内にたまってしまい,ミトコンドリアと呼ばれる細胞内でエネルギーを供給する小器官の機能障害、活性酸素を始めとした細胞にとって毒となる成分の暴露が発病に関与するのではないかと考えられています.
ヘルペスが起こす最も大きな問題は宿主の細胞の核の染色体のゲノムに自分のゲノムを組み込んだ後細胞のDNAを部分特異的組み換えを起こすときに癌関連遺伝子の2種類を突然変異させてがん細胞に仕立て上げてしまうことです。
末梢寛容誘導もあり得ません。末梢寛容誘導というような適応免疫で見逃した寛容を行えるはずもありません。もちろん、バージン T 細胞は、規定の交通パターンから外れて組織に侵入するものもありますどころかバージン T 細胞の、規定の交通パターンは存在しません。誰が規定の交通パターンは決めたんでしょうか?バージン T 細胞は自分のTCRレセプターに合うAPCに出会うためになりふり構わずに必死になって合うAPCを探しまわっているのに規定の交通パターンなんかあるわけはないでしょうに。APCを探すためにはバージン T 細胞の規定の交通パターンで規制をかけられ続ければ永遠にT 細胞はバージンで胸腺で生まれた意味もなくなります。潜在的に自己反応性を持つ T 細胞は、すべての正常な人間の組織に存在しますのも嘘です。一体、潜在的にという意味は何を意味しているのでしょうか?これらの「法を破るバージン T 細胞」は、胸腺では削除を誘発するには稀であるものの、組織ではこれらの T 細胞を活性化するのに十分な自己抗原に遭遇する可能性がありますのも嘘です。この状況に対処するために、自己免疫に対する別のレベルの保護があります。末梢寛容誘導ですなんかあるはずはないのです。
T 細胞の活性化には 2 つの重要な要件があるため、バージン T 細胞は、T 細胞受容体をクラスター化(寄せ集めること)するのに十分な提示された抗原に遭遇するだけでなく、抗原を提示している細胞から共刺激シグナルも受信する必要があります。ここで、活性化された抗原提示細胞(APC)が登場します。これらの特別な細胞は、抗原を提示するための MHC 分子を表面に多数持っており、B7 などの共刺激分子も発現します。対照的に、心臓細胞や腎臓細胞などの通常の細胞は、一般的に MHC タンパク質を高レベルで発現しないか、共刺激分子を発現しないか、またはその両方です。その結果、腎臓抗原を認識する受容体を持つバージン T 細胞は、おそらく腎臓細胞に直接到達しても、活性化されません。バージン T 細胞が細胞上に提示された同族抗原(合う抗原)を認識しても、必要な共刺激を受けない場合、その T 細胞は「無力化」されます。T 細胞のように見えますが、機能しないT 細胞になります。免疫学者は、その細胞は無力化されていると言います。多くの場合、無力化された細胞は最終的に死滅するため、末梢寛容の誘導は無力化または死につながる可能性があります。その結果、T 細胞の活性化中に 2 番目の共刺激「キー」が必要になることで、通常の移動パターンから外れたバージン T 細胞から保護されますというのも嘘です。通常の移動パターンから外れようが外れまいがどうでもいいのです。
活性化による誘導細胞死による寛容もありません。それでは、自己免疫性T 細胞が胸腺での除去を逃れ、交通規則を破り、組織に侵入したらどうなるでしょうか。もともと自己免疫性T 細胞なんか存在しないのでこんな議論は無意味です。そして、この T 細胞が、たまたま T 細胞を活性化するために必要な共刺激を提供できる細胞上の受容体を架橋するのに十分な密度で MHC 分子に表示された同族抗原をたまたま見つけたとします。その場合どうなるでしょうか。すべてが失われるわけではありません。このありそうもない状況で私たちを守ることができる、寛容誘導の「層」がさらにもう 1 つあるからです。このありそうもない状況でも自己面疾患がないのにあるとすれば別の寛容誘導の「層」があるとまで議論したがるのは癌治療に次に自己免疫疾患こそ一番お金が儲かるので言い張り続けるのです。悲しいですね。
前回の講義では、侵入者が倒されたときに T 細胞が排除される 1 つの方法として、活性化誘導細胞死 (AICD) について説明しました。この同じメカニズムは、T 細胞が移動が許されるための交通規則を破り、組織内で自己抗原によって活性化される未処理の T 細胞からも保護するのに役立ちます。このような状況の T 細胞は、常に存在する自己抗原によって何度も刺激を受け、その場合、自己反応性の T 細胞は通常、活性化誘導細胞死によって排除されます。免疫系は、この継続的な再活性化が「自然ではない」と感知し、問題のある自己反応性の T 細胞を排除するかのようです。自己免疫疾患こそ「自然ではない」と何よりも免疫系は知っているでしょうに。
したがって、T 細胞の寛容誘導は多層的なプロセスですというのも嘘です。免疫系は、すべての T 細胞の自己反応性を検査しようとする代わりに、少なくとも 5 つの寛容誘導メカニズムを採用しますのもあり得ない話です。この多層的なアプローチにより、ほとんどの人間では自己免疫疾患が発生しないことが保証されます。ない病気が発生しないのは当たり前のことです。5 つの寛容誘導メカニズムなんか100%存在しません。すべて捏造です。悲しいですね。
長い自己免疫論者の矛盾に満ちたインチキな彼らのいう上の5つの寛容誘導メカニズムを、つまり自己の成分に対してT細胞が非自己として攻撃仕掛けることがないようになる寛容メカニズムが何かを復習しながらまとめてみましょう。存在しないメカニズムを解説するのは不愉快極まりのないことですが頭のよう医学者の間違いを正確に知っておくのも免疫学の復習になります。①中枢寛容②制限されたT細胞の移動経路③二次リンパ器官の抑制④無くなったシグナル⑤活性化誘導アポトーシス
①中枢寛容。中枢の胸腺において胸腺に大量にある自己抗原を認識するT細胞を除去することです。自己抗原は元々何一つありません。
②制限されたT細胞の移動経路。バージンT細胞は組織にある大量の自己抗原は見えません。元々ないものは見えないのです。何故ならばバージンT細胞は組織にでて異動する経路が制限されているからです。制限はされてはいません。
③二次リンパ器官の活性化抑制。二次リンパ器官の活性は自然制御性T細胞によって抑制されているからです。この自然制御性T細胞の正確な作用はいまだはっきりしません。ヘルペスとの戦いは細胞を傷つけ細胞を減らすだけでヘルペスを根絶は絶対にできないのでヘルペスとの共存を目指してヘルペスとの戦いを抑制するために生まれたのが自然制御性T細胞なのです。自然制御性T細胞とは別に誘導性制御性T細胞もありますが殺せない化学物質(アレルゲン)と戦っても化学物質(アレルゲン)は溜まるばかりですからあるどっちみち無駄な戦いですからアレルゲンとは免疫寛容を行って共存するために働くのが誘導性制御性T細胞(iTreg)です。
④無くなった(行方不明の)シグナル。TCRの架橋と補助刺激が無くなったのでバージンT細胞は組織で死んでしまうかアネルギーになってしまう。アネルギーとは、またはアナジーとは、異物に対する生体の防御機構による応答の欠如を示す。抗原が与えられても抗原抗体反応が起こらない状態。アレルギーに対する言葉です。自己の成分は非自己ではないので異物にはならないのでもともとアネルギーであったのです。アッハッハ
⑤活性化誘導アポトーシス。人体を構成している無限大に存在する自己抗原によって慢性的にT細胞は刺激されているのでアポトーシスで除去されてしまうことです。ハチャメチャな論理です。
以上この世に論理的にも実際にも存在しない自己免疫疾患が存在するというバカげたウソの論理がまとめたものです。このようにお金を稼ぐために治療と称して治らない病気でと愚かな医学に無知な大衆をたぶらかす病名ばかりが大手を振っています。ワクチンと抗生物質が完成された現代において最後に残る病気の原因はワクチンが意味を持たないヘルペスだけしか残されていないのです。現代に見られる5大難病といわれる①自己免疫疾患と②アレルギー疾患と③遺伝子病と④原因不明な病気と最後は⑤癌5種類の病気ですがこれらの病気の原因はすべてヘルペスウイルスなのです。ヘルペスについてはここを読んでください。
自己成分に対してB 細胞寛容は必要ではなく存在もしないのですがこの論文の著者は相変わらず「ある」と言い張っています。免疫学者はかつて、受容体が自己抗原を認識する B 細胞を削除する必要はないかもしれないと考えていましたがこれは正しかったのです。潜在的に自己反応性の B 細胞を活性化するために必要な T 細胞は、すでに殺されているか、無力化されているだろうという考えでした。その結果、B 細胞寛容は T 細胞寛容によって「カバー」される可能性があります。しかし、自己反応性の可能性がある B 細胞を寛容化するメカニズムも存在することが明らかになっています。B 細胞とT 細胞は全く違った役割がありB 細胞寛容は T 細胞寛容によって「カバー」されることはできません。
ほとんどの B 細胞は、生まれた場所、つまり骨髄で寛容化されるのは嘘です。そのメカニズムについては誰も知りません。骨髄では生まれたナイーブB細胞は抗体を作るための教育も長いプロセスを時間をかけて末梢の二次リンパ器官でやっと教育され活性化されるのに自己と非自己とを見分ける教育が骨髄でなされることはできないし必要もありません。B 細胞が骨髄で遭遇する抗原は、ほぼすべて自己抗原ですといわれるのですが自己抗原もそもそも存在しないのです。末梢であろうが中枢であろうが免疫システムには見分ける必要もない自己抗原などはどこにも存在しないのです。
自己寛容のB 細胞寛容というこのテストは、T 細胞の胸腺寛容誘導とほぼ同等も嘘です。。B 細胞が遺伝子セグメントを混合して組み合わせ、受容体の遺伝子を構築した後、これらの受容体が骨髄に存在する抗原を認識するかどうかがテストされるシステムはありません。受容体が自己抗原を認識する場合、B 細胞には、軽鎖遺伝子を再編成して、自己抗原に結合しない新しい受容体を作り出す別の機会が与えられます。このテストはあくまでも軽鎖遺伝子にのっとって正しいγさやκさができたかをするためで許容できる受容体を生成して合格するのはすべての B 細胞の約 10% のみです。残りの不完全な軽鎖や重鎖であるBCRを持ったB細胞は骨髄で死滅します。
二次リンパ器官の胚中心における B 細胞寛容の維持はあり得ないという次の文章は久しぶりの正しい唯一の説明ですのでむつかしいのですが読んでおいてください。
胸腺で検査されたときに発現する同じ受容体にとどまる T 細胞とは対照的に、B 細胞は二次リンパ器官で活性化された後、体細胞超変異によって受容体を変更する可能性があるのは当然です。したがって、体細胞超変異を起こしている B 細胞が自己抗原を認識できる受容体を持つようになるのではないかと疑問に思うのは自己免疫論者だけです。もしそうなら、これらの B 細胞は自己免疫疾患を引き起こす可能性のある抗体を生成する可能性があります。幸いなことに、これは幸いなことに発生しないことが判明していなくても真実です。その理由は次のとおりです。B 細胞が自己抗原を認識できる受容体を持つようになることは無いと自分が言っているのに幸いなことに常発生しないことが判明しているのに何故説明するのでしょうか?正しい二次リンパ器官の胚中心における B 細胞寛容の維持はあり得ないという文章を伝えたいためです。珍しいですね。
B 細胞が胚中心で hyper‐mutation(体細胞超突然変異) し、その受容体が自己抗原を認識するようになった場合、濾胞樹状細胞で宣伝されている自己抗原を見つけて刺激を受ける可能性は非常に低くなります。結局のところ、FDC はオプソニン化された抗原のみを表示します。そして、自己抗原はオプソニン化されません。何故ならばオプソニン化は自然免疫の役割ですから自己の安全な分子に対してはオプソニン化できないのです。オプソニン化できるのは病原体の非自己である人体に有害な分子に対してだけです。だから安全な自分を傷つけることがない自己の成分を免疫が攻撃することはつまり自己免疫疾患は絶対に起こらないのです。したがって、潜在的に自己反応性の B 細胞が胚中心で直面する最初の困難は、濾胞樹状細胞上のオプソニン化された自己抗原の欠如です。したがって二次リンパ器官の胚中心における B 細胞寛容の維持は不可能というべきです。自己免疫はしかし、共刺激の欠如という別の問題がさらにあるのでますますB 細胞寛容の維持はありえないのです。
濾胞ヘルパー T 細胞(Tfh)は、二次リンパ器官の T 細胞領域で活性化された後、リンパ濾胞に移動して B 細胞を支援します。この支援は、濾胞ヘルパー T 細胞 (Tfh 細胞) と B 細胞が互いに刺激し合うダンス中に行われます。このダンスに参加する B 細胞は、活性化されたAPCでありB細胞受容体(BCR)が結合する抗原を内部化しておりTLRを持っており、クラス II MHC 分子を使用してこの抗原を濾胞ヘルパー T 細胞( Tfh )に提示します。B 細胞はAPCであるので、Tfh 細胞がアクティブであるために必要な共刺激 ( B7) も提供します。その見返りとして、Tfh 細胞は B 細胞に必要な共刺激 ( CD40L) を提供します。ここで重要な点は、この双方向刺激が機能するためには、Tfh 細胞の受容体と B 細胞の受容体が同じ抗原、より正確には、同じ抗原の一部(エピトープ)を認識する必要があるということです。B 細胞が体細胞超突然変異(hyper‐mutation))を起こし、その BCR が自己抗原に結合することもできないし、内部化し敵の分子とも認識できるTLRと結合することはできないので、自己抗原を提示することはできないので、その新しい抗原は「必要な」Tfh 細胞の受容体によって認識されません。その結果、B 細胞と T 細胞は協力して互いを刺激することができなくなります。両者は「共通の関心」を失ってしまいます。また、B 細胞は胚中心で生き残るために Tfh 細胞の助けを必要とするため、B 細胞と Tfh 細胞の相互依存性により、B 細胞は体細胞超突然変異を起こしても「軌道」に乗ったままです。そのため、B 細胞の超突然変異中に自己寛容性が維持されるのは、効率的な BCR シグナル伝達に必要なオプソニン化された自己抗原がないことと、自己抗原を認識する B 細胞を助けることができる胚中心 Tfh 細胞がないことの 2 つの理由により二次リンパ器官の胚中心における B 細胞寛容の維持はあり得ないのです。こんなに正直に真実を語ることができる免疫学者は自己免疫疾患でお金を全世界の医者が儲けるために真実の事実よりもあり得ない論理を展開して「と思われる」とか「と考えられる」とかの誤った空想を書きまくるのか理解に苦しみます。しかもこの論文を書いている免疫医学者はニュウーヨークにあるコロンビア大学の著名な教授であった人ですから自然免疫も適応免疫のすべてに通じている人なのに「治療学」の話しになると一から十までウソになるのは何故でしょうか?彼が隠している真実である
自然免疫のナチュラル キラー細胞の正の選択もあり得ません。ヘヘルペスウイルスを含む多くのウイルスは、感染した細胞上のクラス ⅠMHC 分子の発現を低下させることで免疫システムを回避しようとします。この卑劣な策略は、キラー T 細胞がこれらの細胞を「調査して」感染していると判断するのを防ぐように設計するのに最も秀でているのはヘルペスウイルスです。このヘルペスウイルスの策略に対抗するために、ナチュラル キラー細胞は接触した細胞を調査し、表面にクラス Ⅰ MHC 分子を表示していない細胞を破壊します。これは、敵を認識するには自己である自己だけが持っているクラス Ⅰ MHC 分子の上に敵の成分を乗せて初めて敵を認識できるからです。これが機能するのは、NK 細胞が表面に「抑制性受容体」を持っているためです。この受容体は健康な細胞上のクラス ⅠMHC 分子を認識し、「殺さないで」という信号を伝えて、自己の身分証明となる分子を持っているのは自分の細胞を殺さないようにする印です。しかし、MHC 分子は非常に遺伝子の多形性があります。つまり、各 MHC 遺伝子が発現されると、ヒト集団内でわずかに異なる多くのMHC分子の形態を持っているので、私の NK 細胞上の抑制性受容体が私自身の少し異なった蛋白質のクラス ⅠMHC 分子を認識しない可能性があります。そして、NK 細胞上の抑制性受容体が自分自身のクラス ⅠMHC 分子を認識できなければ、私の NK 細胞は、すべての正常細胞がウイルスに感染していると考え、それらを殺してしまうかもしれません。この問題の解決策の 1 つは、各クラス I MHC 分子をそれに対応する NK細胞の殺すなという 阻害性受容体と共発現させることですが、実際にはそうではありません。むしろ、すべての人間は阻害性 NK 受容体の遺伝子を複数持っており、これらの遺伝子も非常に多型性があります。その結果、すべての人が阻害性受容体遺伝子の集合を受け継いでおり、これらは通常、人によって異なります。さらに、これらの遺伝子は発現のためにランダムに選択されるように見えるため、異なる阻害性受容体の配列は、同じ人の中でも細胞ごとに異なります。
現在の考え方では、NK 細胞が完全に機能して細胞を殺すには、「殺傷する許可」を得る必要があると考えられています。この許可を得るには、私たちの NK 細胞は、細胞上のクラス I MHC 分子の少なくとも 1 つを認識できる阻害性受容体を持っている必要があります。自己 MHC 分子に結合する阻害受容体を持たない NK 細胞は、機能できないように無力化されます。その結果、NK 細胞は NK 細胞媒介自己免疫を回避するためにスクリーニングされるといわれていますがNK 細胞はどのようにして自己寛容性についてスクリーニングされ、この「検査」は体のどこで行われるのでしょうかなどは誰にもわかりません。私たちの免疫システムがどのように機能するかについては、まだ解明されていないことがたくさんありますが自己免疫疾患の有無もそのひとつですが堂々と存在する病気として治療が続けられています。本当はヘルペスによる感染症に過ぎないにもかかわらず。残念です。
この賢すぎますが金儲けのために頭が狂った学者の最後のまとめです。この講義では、免疫学における最も重要な謎の 1 つであるどころか偽りの、同じ T 細胞受容体(TCR)がどのようにして正の選択 (MHC 制限)、負の選択 (寛容誘導)、および活性化を媒介するのかについて説明しましたが負の選択 (寛容誘導)はないのです。現在の考え方では、胸腺における T 細胞の正の選択 (生存) は、細胞の TCR と皮質胸腺上皮細胞に表示された MHC 自己ペプチドとの間の比較的弱い相互作用の結果であると考えられていますがこの文章の中の「自分のMHC と自己ペプチドとの間の比較的弱い相互作用の結果である」の「比較的弱い相互作用言の文章に中の「比較的弱い」という言辞は科学である免疫学において使われるべきではないのです。何故ならば「比較的」も「弱い」という言葉はあまりにも文学的で科学や医学の明晰な表現ではない言葉であるからです。この曖昧な「テスト」は、MHC 分子によって提示された抗原に T 細胞の注意を集中させ、認識が「ネイティブ」抗原ではなく提示された抗原に限定されるようにすることを目的としていますが曖昧過ぎます。
胸腺における T 細胞の負の選択 (死) は、アポトーシスではなく細胞の TCR と髄質胸腺上皮細胞(mTEC)または胸腺樹状細胞(TDC)に発現された MHC 自己ペプチドとの間の強力な相互作用によって引き起こされます。T 細胞の負の選択 (死) は髄質胸腺上皮細胞(mTEC)または胸腺樹状細胞(TDC)の強力な相互作用はどんな相互作用なのかは一切詳しくは書かれていません。この「試験」は、自己免疫疾患を引き起こす可能性のある T 細胞を排除するように設計されています。この設計図がどんなものか知りたいのですが初めて耳にした遺伝子の設計図なのですから詳しく書いてもらいたいのですが説明は一切ありません。最後に、胸腺を離れた後、T 細胞は活性化され、その TCR と専門の抗原提示細胞によって表示されるMHC―ペプチド複合体との強力な相互作用を通じて、病気から私たちを守ることができますという働きが本来のT 細胞の機能であって、T 細胞は自己免疫疾患には全く関りがないのです。
胸腺 (中枢) 寛容誘導はかなり優れていますが、完璧ではありませんのは胸腺 (中枢) 寛容誘導も存在しないからです。人間の知恵は、生命の進化が作った神なる働きは「かなり優れていますが、完璧ではない」のはあり得ないのです。C方法の 1 つは、血液、リンパ、および二次リンパ器官へのバージン T 細胞の輸送を制限することですが、血液、リンパ、および二次リンパ器官へのバージン T 細胞の輸送はすべて多かれ少なかれつながりがあるので血液、リンパ、および二次リンパ器官へのバージン T 細胞の輸送を誓言する必要もないうえに制限すれば免疫に司令官であるバージン T 細胞は活性化されて本来のサイトカイン工場でもある機能が果たせなくなります。又二次リンパ器官に豊富に存在する抗原を認識する受容体を持つ T 細胞は、同じ抗原も豊富に存在する胸腺で効率的に削除されますといわれますが果たして胸腺という組織に存在する限られた細胞が持つ自己抗原が何種類あると思いますか?更にT 細胞が認識する多種類の同じ自己抗原も豊富に存在していると思いますか?胸腺に住んでいる自己の細胞は極めて少ないのですよ?逆に、胸腺で自己反応性 T 細胞が削除を逃れるほどに希少な自己抗原は、二次リンパ器官のバージン T 細胞を活性化するには希少すぎるのが普通です。したがって、バージン T 細胞は輸送パターンが制限されているため、胸腺で希少な自己抗原を機能的に認識しないままです。二次リンパ器官には一次リンパ器官である胸腺と同じく自己の細胞が限られているので二次リンパ器官の天然制御性 T 細胞(nTreg)も、自己免疫に対する保護を提供するのは実は胸腺と同じなのです。また、バージン T 細胞が血液 – リンパ – 二次リンパ器官系の外に出た場合、活性化ではなく、アネルギーまたは死につながる状況で自己抗原に遭遇します。バージン T 細胞が活性化されず前に死ぬことは聞いたことがありません。さらに、組織内の自己抗原を認識することで活性化されるといわれるこの世に絶対に存在しないこれらのまれな古くなってしまった過去の T 細胞は、通常、慢性の再刺激によって死にます。これを活性化誘導細胞死(AICD)といいますがあくまでも非自己に対する敵に対して当てはまる用語です。アネルギーとはリンパ球が特異的な抗原に応答しない場合は、アネルギー性があると言われていますが逆にアネルギー細胞が持つ特徴は、健康人の体内に自己に反応するCD8陽性T細胞がアネルギー状態で存在すると、自己免疫応答を抑制しているのですといわれますが自己成分を認識するT細胞は存在しませんからどちらも間違いです。
T 細胞には、中枢寛容が誘導される胸腺という別の器官がありますが、豊富な自己抗原を認識する受容体を持つ B 細胞はあり得ないのですが、生まれた場所、つまり骨髄で排除されます。このスクリーニング中に、自己反応性 B 細胞には、自己抗原を認識しない BCR を作り出すために、受容体を「編集」する 2 度目のチャンスが与えられますといわれるのも嘘です。
T 細胞と同様に、B 細胞における寛容誘導は多層的です。バージン B 細胞は、主に血液、リンパ、二次リンパ器官を通過します。したがって、T 細胞と同様に、ナイーブ B 細胞のトラフィック パターンは、通常、骨髄での寛容誘導中にテストされなかった豊富な自己抗原との接触からそれらを保護します。血液/リンパのトラフィック パターンから外れたナイーブ B 細胞は、通常、BCR を架橋できる形態の十分な自己抗原に遭遇しません。さらに、組織内の自己抗原によって受容体が架橋されたバージン B 細胞は、通常、活性化に必要な共刺激シグナルを受信しません。また、共刺激なしで架橋すると、B 細胞がアネルギー化または死滅する可能性があります。いずれも嘘ばかりです。
B 細胞が胚中心で成熟すると、体細胞超変異(hyper mutation)を起こして受容体の親和性を高めることができます。このプロセスにより、変異した BCR が自己抗原を認識する可能性はゼロです。幸いなことに、これは通常問題になりません。B 細胞が胚中心で増殖するには、その受容体が濾胞樹状細胞によって表示されるオプソニン化された抗原を認識する必要がありますが、自己抗原は通常オプソニン化されません。さらに重要なのは、胚中心の濾胞ヘルパー T 細胞は、変異した BCR が認識して提示する自己抗原を認識しないことです。そして、B 細胞は生存のために Tfh 細胞の助けを頼りにしています。
B 細胞または T 細胞を寛容化するメカニズムはどれも絶対確実ではないのはあり得ない現象が存在するという仮定から話が始まるからです。自己反応性の可能性のある細胞を捕らえるための寛容誘導メカニズムが複数層あるため、システム全体は非常にうまく機能し、深刻な自己免疫疾患に苦しむ人は比較的少数なのはヘルペスが増えすぎたためにてウソの自己免疫疾患の症状が出現する人は少ないからです。
ナチュラル キラー細胞も自己反応を回避するためにテストされていますといわれますがこれも嘘です。 NK 細胞に、人のクラス I MHC 分子の少なくとも 1 つを認識する抑制性受容体がない場合は果たしてあり得るでしょうか?その NK 細胞は機能しなくなります。このプロセスにより、NK 細胞が健康な細胞を攻撃して自己免疫を引き起こすことが防止されますというのは本来そんなNK 細胞は生まれません。。
誤った免疫システムというサブタイトルは間違いです。というのは免疫システムは免疫の遺伝子に人体を病原体から守るために36億年かけてダーウインが説いているように生きるために環境に適応するために進化し遺伝子に刻まれているからです。適応免疫もその進化の所産の一つです。自分を傷つけるなどを行う免疫システムを持った種は即滅びてしまっているからです。生き続けることができないからです。嘘をついてお金を儲けようとする小賢しい狡賢い医学者がディープフェイクで捏造した病気が自己免疫疾患です。彼らは免疫学の理論を駆使して嘘ばかりつきまくっています。その嘘を暴露するために私はこの論文を書き続けているのです。病気を作り続けなければ医薬会は崩壊してしまうからです。人間は自分の快楽を得るために生きているのです。他人が病気で苦しもうがどうでもいいのです。残念です。医療法は病気を治して初めて医療報酬を医者が得られるように改正すべきですがそうすれば医者という職業はなくなってしまうでしょう。
自己免疫疾患はすでに何回も証明したように自己である成分を絶対に攻撃できないのです。
ヒトの免疫システムは、B細胞やT細胞でさえも自己に対する寛容性を注意深くチェックするような完璧なシステムのために、膨大な生物学的「エネルギー」を費やしているわけではない。その代わりに、このシステムはさらに膨大なエネルギーを過当なほかの多層的な戦略に頼っているというのも嘘なのです。適応免疫の最高峰にいるB細胞やT細胞よりも進化のレベルにおいてはるかに下等な各層には、自己反応細胞を淘汰する力も人体を自己を病原体から守る力B細胞やT細胞よりもはるかに劣っているのにどうしてそんな自己と非自己とを見分け選択する高等な機能を持ち合わせていると思いますか?
上の層での寛容誘導をすり抜けた細胞を下の層がキャッチする。これは通常、非常にうまく機能するのだが、時折「ミスを犯す」ことがあり、外敵から私たちを守る代わりに、免疫システムの武器が私たちに反旗を翻してしまうのである。下層なレベルの免疫機能がどのようなミスをどのように起こすのか説明は一切されていません。金を儲けるためにとにかくまず「自己免疫疾患ありき」から始めているので論理が支離滅裂なのです。自己免疫疾患は、自己に対する寛容を維持するためのメカニズムが、病的な状態を引き起こすほど深刻に破壊されたときに生じる根拠も何一つありません。。アメリカ人のおよそ5%が何らかの自己免疫疾患に苦しんでいるのは資本主義の大国である対抗であるアメリカほど金持ちと貧乏人の賃金格差が大きい国は他にありません。格差を示す時に係数は0から1までありますが0,48で暴動が起こってもよいくらいになっていますがアメリカの下層階級の人たちはストレスに耐えに耐えてヘルペスを増やして自己免疫疾患で苦しんでいるのですが医者たちはない病気つくりに励むばかりです。悔しいですね。
自己免疫の中には、遺伝的欠陥に起因するものもあるのも嘘です。。例えば、ほとんどの自己免疫疾患は、自己反応性リンパ球が繰り返し刺激される慢性疾患であるのも嘘です。健常人では、慢性的に刺激されたT細胞が、その表面にあるFasタンパク質がFasと結びつくことによって排除される自己免疫疾患を起こす活性化誘導性細胞が死ぬことによって、この活性化が制御され自己免疫疾患が起こらないので嘘です。FasまたはFasリガンドタンパク質のいずれかに遺伝的欠損を持つヒトは、この寛容防御の層を欠くため、自己抗原によって慢性的に刺激されてもT細胞は死滅しない。とうとう自己免疫疾患を遺伝子業に仕立て上げてしまいました。その結果生じる病気、自己免疫性リンパ増殖症候群またはカナレ・スミス症候群は、その病理学的結果として、リンパ節の巨大な腫脹、自己抗原を認識する抗体の産生、二次リンパ器官への多数のT細胞の蓄積をもたらすのも膨大なヘルペスが増えすぎて起こっているだけで遺伝子疾患でもないのです。Fasとは何でしょうか? Fasリガンドタンパク質とは何でしょうか?Fasは Fas受容体(FasR)であり細胞表面に位置するsaibouwo細胞死なせる死受容体であり、そのリガンドであるFasリガンド(FasL)が結合した場合にプログラム細胞死(アポトーシス)をもたらす。この経路は2つのアポトーシス経路のうちの1つであり、もう1つはミトコンドリアを介した経路です。
Fasリガンドは、膜貫通型蛋白質であり、腫瘍壊死因子ファミリーの一員である。アポトーシスを誘導するFas受容体に結合する。FasリガンドとFas受容体相互作用は、免疫系の制御や悪性腫瘍の進行において重要な役割を果たしている。
Fasリガンドは細胞表面受容体Fasを介し, 細胞表面受容体Fasを持っている標的細胞にアポトーシスを引き起こすサイトカインである。FasはTNF/NGF受容体ファミリーに属するI型膜貫通蛋白質で,様々な細胞に広範に発現している. NGF受容体とは神経成長因子受容体です。一方,FasリガンドはTNFファミリーに属する膜貫通蛋白質として産生される。Fasリガンドは主に活性化T細胞やNK細胞に発現し,細胞傷害因子として働いている.また,免疫隔絶組織として知られる精巣や目の非造血系の細胞にもFasリガンドが発現しており,このことがこれらの組織が免疫系の攻撃を受けない原因になっているのです。
一部の自己免疫疾患は遺伝的欠陥によって引き起こされるが、自己免疫疾患の大部分は、寛容誘導機構の層が遺伝的に正常な人の自己反応細胞を排除できないために起こるといわれているが遺伝的欠陥であれば生まれた時から自己免疫疾患があるはずです。
最新の考えでは、考えであって真実ではないのですよ。自己免疫が起こるためには、少なくとも3つの条件が満たされなければならない。第一に、標的となるT細胞が認識不能な自己抗原に由来するペプチドを効率的に提示するMHC分子を発現していなければならない。
自己免疫の第二の条件は、罹患者が自己抗原を認識するレセプターを持つT細胞、場合によってはB細胞を産生することである。TCRとBCRはミックス・アンド・マッチ戦略(遺伝子の組み換え)で作られるため、ある個人が発現するレセプターのレパートリーは他のすべてのヒトのレセプターとは異なり、リンパ球が死滅して入れ替わるにつれて時間とともに変化するのは間違いです。何故ならば造血幹細胞の遺伝子が変わらない限りあり得ないことです。一卵性双生児が発現するTCRやBCRのコレクションでさえも異なるでしょう。TCRやBCRのコレクションでさえも異なるのはヘルペス感染によるものですが、病気ではありません。したがって、ある人が特定の自己抗原を認識するレセプターを持つリンパ球を産生するのは、ほとんど偶然の産物なのですがそれが直接、自己免疫疾患の原因とはなりません。何故ならば受精卵の精子と卵子の遺伝子の組み合わせは100%偶然そのものであるからです。
つまり、自己免疫疾患が起こるためには、自己抗原を提示できるMHC分子と、自己抗原を認識できるレセプターを持つリンパ球がなければならない。しかし、これだけでは十分ではない。自己反応性リンパ球を排除するように設計された寛容機構を崩壊させる環境因子も必要である。何年もの間、医師たちは自己免疫疾患が細菌やヘルペスウイルスの感染に引き続いて起こることが多いことに注目してきたと書かれていますが、ヘルペスウイルスの感染が自己免疫疾患であるのに世界中の偉い医学者は誰も気が付いていないだけです。そして免疫学者たちは、微生物の攻撃は自己免疫疾患を引き起こす重要な環境因子の一つではないかと考えているのですが微生物であるヘルペスウイルスは自己免疫疾患その病気自身を引き起こしているのです。ヘルペスウイルスや細菌感染がすべてではないことは明らかであるという言い方も間違っているのです。自己免疫疾患には細菌感染は全く関係がないのですがヘルペスウイルス感染は自己免疫疾患のみならずすべての癌の原因である800個以上もあるゲノムの癌関連遺伝子を突然変異させて癌化させているだけなのです。遺伝的素因(例えば、受け継いだMHC分子のタイプ)や自己反応性レセプターを潜在的に持つリンパ球との組み合わせで、微生物感染は自己免疫疾患を引き起こす「最後の藁」になるかもしれないといわれますが正に自己免疫疾患はヘルペスと免疫の戦いの症状にすぎないのです。
分子模倣については
感染症が自己寛容の崩壊につながる理由を説明するために、免疫学者が現在最も気に入っている仮説は、分子模倣と呼ばれるものである。80年前にワクチンと抗生物質分子で病原体で人類の誕生以来、苦しんでき感染症で死ぬことは無くなったので医療でお金を稼ぐためにイギリスの超有名な大学の医学部から自己免疫疾患が生まれました。この世にない自己免疫疾患が何故お金が患者が死ぬまで稼げるかの理由は患者が生きている限り自分の成分を免疫が攻撃し続けると死ぬよとウソをついた論理はユダヤ思考そのものです。大衆は医学についてはバカですからヘルペスが原因である症状がみられる病気のすべてを自己免疫疾患に祭り上げたのです。
免疫システムは何の目的のために生まれどの様に生きるためになければ人類は存続不可能であるのにその免疫が人体を攻撃して死ぬことがあるなどという発想は正に頭の良い金儲けしかないユダヤ的発想です。
本来の非自己である病原体を攻撃する非自己的免疫と守るべき自己を攻撃するウソの自己免疫疾患の違いは何でしょうか?現代においては、私たちの身の回りには、細菌やウィルス、など生体に悪影響を起こす生きた病原体と死んだ花粉、粉塵に含まれている化学物質という異物に生物や物質(外敵)に満ち溢れていますが、ワクチンと抗生物質があるのでワクチンが作れないヘルペスウイルス以外のすべての病原体を気にせず生きていくことができます。化学物質に対しては免疫寛容によって共存できます。
一般に使われている免疫という言葉は、獲得免疫系のTリンパ球を指す場合が多く、外敵(抗原)が侵入すると、その抗原に対応する抗体を産生する液性免疫と細胞内に感染した病原体であるウイルスは細胞内に感染します。とりわけワクチンが効果のない最も多いヘルペスウイルスは細胞性免疫で細胞もろともヘルペスウイルスを殺します。最後に残った病原体であるヘルペスが自己免疫疾患を作らない免疫の働きの概要を説明します。
生体にヘルペスが侵入すると三つの抗原提示細胞であるマクロファージと樹状細胞とB細胞が抗原を捕捉して取り込んだ抗原を分解し活性化した抗原提示細胞抗原はこのヘルペスという外来の非自己であるに特異的なヘルペスの情報である人間が持っていないヘルペスのペプチドだけをMHCを結び付けてヘルパーT細胞に提示します。 するとこの活性化したヘルパー T細胞がさらに活性化してヘルペスの抗原情報をB細胞とTCR(T‐cell receptor)とBCR(T‐cell receptor)を介してヘルパー T細胞とB細胞結合しての活性化して同じ敵を認識したT細胞の指示を受けて特定の抗体産生細胞に変化して同じ敵を認識したB細胞は形質細胞になって抗体を産生します。一方同じ敵を認識したキラーT細胞の活性化は同じ敵を認識した活性化したヘルパー T細胞を介し殺す能力が発揮されるのです。
自己免疫疾患は、上で述べたように、三つの抗原提示細胞であるマクロファージと樹状細胞とB細胞自己の生体成分を認識できないのでヘルパー T細胞とB細胞に提示できないのでT細胞とB細胞は自分の成分のペプチドに対しては認識することができないので自己の成分を敵に回すことは絶対に不可能であるように自然免疫のレベルの抗原提示細胞の敵を見分ける方法を進化の中で遺伝子に刻印されてしまったのです。当たり前のことなのです。というのはマクロファージは生きた人間の成分を食べてしまったらどうなるでしょうか?とすれば人食いマクロファージが高等免疫が生まれる前に「人食いマクロファージ」によってとっくの昔に絶滅していたことでしょう。長い免疫のス¥以前免疫のレベルでや細胞を「異物」と誤認識することにより、自己の細胞や組織を攻撃してしまう病気ですがなぜ誤認識されるのかについて詳細はわかっていませんが、「自己」か「異物」かの判断は主にT細胞に委ねられています。抗原提示細胞では、取り込んだ抗原をペプチドにまで分解しT細胞に提示しますが、提示される抗原ペプチドす。
自己免疫疾患にはどのような病気があるの?
ウソの自己免疫疾患を起こす場合、ヘルペスがどの細胞や組織などに感染するかによって病状が異なります。例えば、中枢神経の髄鞘の細胞に感染すると、神経の髄鞘が攻撃され神経系統が破壊され、多発性硬化症という病気が発症します。ヘルペスウイルスはあらゆる細胞に感染する特性を持っていますので一般に、自己免疫疾患は、全身にわたる細胞にヘルペス感染が起こると影響がおよぶ全身性自己免疫疾患と、特定の臓器だけにヘルペスが感染するとヘルペスと免疫との戦いによる炎症の影響を受ける臓器特異的疾患に分けられます。全身性自己免疫疾患には、関節リウマチや全身性エリテマトーデスなどの膠原病があります。膠原病も結合組織病もすべて原因はヘルペスであり病名が異なるだけですから自己面疾患とつけようがどうで見いいことです。バカな医者どもの趣味は「病気つくり」と「病名作り」と病気を治す責任を果たして稼ぐべきお金を儲けることです。アッハッハ!!!!臓器特異的疾患には、多発性硬化症や最近増加してきている潰瘍性大腸炎の他、強皮症、ベーチェット病、突発性血小板減少性紫斑症、重症筋無力症など数多くの難病があります。難病の定義は「一生治らない病気」ですからウソのあり得ない自己免疫疾患のすべてが難病です。下の次の表は、難病情報センター特定疾患医療受給証交付件数のうち自己免疫疾患について1974年から2007年までの交付件数を示したものですが、いずれの疾患も年々増加していることを示しています。自己免疫疾患のすべてが難病ですから特定疾患医療受給証交付はすべての自己免疫疾患の患者さんに与えるべきです。特定疾患医療受給証の意味は実は病気を作ってごめんなさい。でもただの病気つくりでは申し訳ないのでお金を払わなくてもよい病気つくりですよというメッセージなのです。ワッハハッハ!!!!!!
欧州リウマチ学会(EULAR 2023、5月31日~6月3日)で「19種の自己免疫疾患全体の人口に対する有病率は約10%で、患者数は過去20年間に4%増加していた」と発表。人口の1割が自己免疫疾患とは恐ろしい数ですよ。悲しいですね。人口の1割が医者にお金を取られて一生苦しんで生きなければならないのですよ!!!!
さらに恐ろしい殺しの文章が続くのでそれを掲載しておきます。多くの自己免疫疾患は、潜在的に発症し慢性的に経過してやがて自覚症状が出て初めて病院に行き疾患として診断され治療される、といった経過を経ることが多い病気です。そのため、多くの自己免疫疾患の場合、発症してからかなりの時間経過を経て疾患と認識されますので、非常に治りにくく、原因も明確でないことから難病に指定されています。そのため、日本では公費負担の対象として定められている特定疾患に指定されています。また、多くの自己免疫疾患では、女性の患者が多いといわれています。理由は明らかになっていませんがホルモンが関与しているという説もあります。上の数行の文章はバカで無知で真実を嫌悪する医者たちに病気を作って大きな富の作り方を教唆しているのです。
分子模倣による自己免疫疾患が生まれるウソの仮説を紹介しましょう。感染症が自己寛容の崩壊につながる理由を説明するために、免疫学者が現在最も気に入っている100%でたらめな仮説は、分子模倣と呼ばれるものである。
なぜ自己免疫疾患が生まれるウソのパレード仮説が次々と生まれ始めたのでしょうか? 80年前にワクチンと抗生物質分子で病原体で人類の誕生以来、苦しんでき感染症で死ぬことは無くなったので医療でお金を稼ぐためにイギリスの超有名な大学の医学部から自己免疫疾患が生まれました。この世にない自己免疫疾患が何故お金が患者が死ぬまで稼げるかの理由は患者が生きている限り自分の成分を免疫が攻撃し続けるので死ぬよとウソをついた論理はユダヤ思考そのものです。大衆は医学についてはバカですからヘルペスが原因である症状が見られる病気のすべてを自己免疫疾患に祭り上げたのです。今でもヘルペスは皮膚科の病気であると一般大衆は信じています。違うのです。現代残された癌や自己免疫疾患や原因不明な病気や先天的であろうが後天的であろうが遺伝子病などの治らない病気のすべての原因はヘルペスなのです。
ウソの分子模倣の仕組みは以下の通りである。
リンパ球はBCRまたはTCRを持ち、その抗原を認識する。しかし、この抗原が単一であることはほとんどない。一つのMHC分子が同じ全体的特徴(長さ、結合モチーフなど)を持つ多数のペプチドを提示できるように、TCRやBCRは通常、複数の異なる抗原を認識(交差反応)することができる。一般に、TCRやBCRは、これらの認識抗原のうち1つまたはいくつかに対して高い親和性を持ち、他の抗原に対しては低い親和性を持つ。微生物が侵入すると、微生物抗原を認識するレセプターを持つリンパ球があるでしょうが自己の成分であるすべての分子を認識することはできないので実は活性化することはないのです。というのは何回も書きましたが三つの活性化した抗原提示細胞である樹枝状細胞と、大食細胞と膜抗体を持ったBリンパ球の三つが敵と認識してTCRやBCRに提示して反応するのは病原体の分子しかないので自己の分子は提示してもTCRやBCRは知らん顔をするからです。
乾癬と分子模倣と炎症については、私もヘルペスによる乾癬に数十年前からかかっていますがストレスが強いときに皮膚の表皮細胞に感染したヘルペスが増殖してケラチノサイトが崩壊して生じて生まれたヘルペス感染症が尋常性乾癬です。あらゆる自己免疫疾患はヘルペス感染症と言えます。嘘の自己免疫疾患である乾癬は、米国人口の約2%が罹患している。最も顕著な症状は、皮膚表面の肥厚と鱗屑である。最も重篤な症例では、これらの「プラーク」が皮膚面積の10%以上を覆うこともある。最近、この疾患は、高レベルのIL-17(そう、キラーT細胞はサイトカインを産生できるのだ!)を産生するCD8+T細胞によって引き起こされることが発見された。IL-17 を産生する CD4 陽性 T 細胞は,既知の Th1 細胞,Th2 細胞とは異なる集団として同定され,Th17 細胞と命名された2)3).IL-17 受容体はマクロファージなどの免疫細胞,角化細胞を含む上皮細胞,血管内皮細胞,線維芽細胞,滑膜細胞などで発現される4).IL-17 が受容体に結合すると,これらの細胞で炎症性サイトカインやケモカインが産生され,局所で好中球の遊走,組織障害,血管新生が誘導される4).近年,Th17 細胞/IL-17は,乾癬,関節リウマチ,多発性硬化症,クローン病などの慢性炎症性疾患において病因的役割を担っていることが明らかになL-17は皮膚細胞(ケラチノサイト)に結合し、ケラチノサイトを増殖させてプラークを形成させる一連の分子イベントを引き起こすことができます。現在提唱されている 「分子模倣 」モデルは、遺伝的に影響を受けやすい人(例えば、クラスⅠMHC分子の特定のバージョンを持つ人)のT細胞は、特定のケラチンタンパク質と溶連菌(これは時々、世界の人口の大部分に感染する)によって作られるタンパク質の両方を認識するレセプターを持っているというものである。このような溶連菌感染に反応するCTLの一部は、患者のクラスⅠMHC分子が提示するケラチンタンパク質と交差反応するTCRを持っているという仮説がある。証拠が全くない自己面疾患の仮説ばかりでしかも考えられているという空想の話しばかり長い長いインチキ話で飽き飽きしているのですが最後まで頭に良い免疫学者のウソのつき方を勉強していますと言いたいところですが残念ながら真実しか興味がないので100%ウソ話には辟易してしまっているのですが最後まで免疫学を武器にしてどのようにウソをつくのかを知りそのウソのウソたるゆえんを暴くために反論を続けます。
自己面疾患一旦細菌感染によって活性化されると、これらの自己反応性CD8+T細胞はIL-17を産生し、誤ったケラチノサイトの増殖とプラーク形成を引き起こす。クラスⅠMHC分子のこれらの特定のバージョンは、免疫系が連鎖球菌感染から身を守るのに役立つため、進化の過程で選択されたのかもしれない。結局のところ、原始人にとって連鎖球菌感染症は命取りになりかねないが、原始人は肌の見た目などあまり気にしていなかったのだろう!
関節リウマチは、世界人口の約1%が罹患する全身性の自己免疫疾患であるのも100%ウソです。関節のポリモーダル自由神経終末の痛覚受容体に感染したヘルペスが原因なのですが、免疫でだけでは殺し切れないので慢性炎症が特徴であるので抗ヘルペス剤と漢方煎じ薬で治ります。この自己免疫反応の標的のひとつは、ある種の軟骨タンパク質であると推定されていますがウソです。軟骨にも数多くのポリモーダル自由神経終末の痛覚受容体がありここにヘルペスが感染するからです。
IgG抗体のFc領域に結合できるIgM抗体は関節リウマチ患者の関節に多く存在する。Fc領域とは免疫グロブリン(IgG抗体)の重鎖定常領域のことです。これらの抗体はIgM-IgG抗体複合体を形成し、関節内に侵入したマクロファージを活性化し、炎症反応を増大させるといわれていますが、本当にIgG抗体のFc領域に、IgM抗体がどのように結合するかについても一切論理的な説明はありません。関節に炎症を起こしているのは関節の様々な細胞に感染したヘルペスと免疫が戦って炎症が起こっているだけです。現代の病気の最後に残された原因のすべてはヘルペスなのです。ににもかかわらず世界の医学者は免疫を落としてヘルペスを増やして病気を作りまくっていますが、しかも最後はヘルペスが作る癌まで作って患者を殺していますが誰も責任を取りません。悲しいですね。
セリアック病とは何でしょうか?グルテン不耐症とは何でしょうか?セリアック病は、小麦や大麦、ライ麦に含まれるタンパク質のグルテンに対する遺伝性の不耐症であり、小腸の粘膜に特徴的な変化を起こし、吸収不良が生じます。タンパク質のグルテンに対する遺伝性の不耐症であるので食べものの「不耐症」とは、特定の食たべものを消化できないために、それを食たべることに耐えられないという意味です。乳糖を消化できないことで起こります。その理由は、乳糖を消化酵素であるラクターゼが小腸で十分に作られないためです。入党を消化する酵素のラクターゼ不足をもたらす先天的酵素欠乏症です。
最後に、エリテマトーデスは全身性の自己免疫疾患はなくてこの世に自分を攻撃する免疫は存在しないので原因はヘルペスですがアメリカでは約25万人が罹患し、その約90%が女性であります。世界で最もストレスが強いのはアメリカの女性です。この病気は、額や頬の赤い発疹が病名の由来となった「赤い狼」のような外観を与えます。他に肺の炎症、関節炎、腎臓障害、脱毛、麻痺、けいれんなど、複数の症状を示すことがある。狼瘡は、顔の額や頬の毛細血管の内皮細胞にヘルペスが感染したために血管が拡張して赤くなっただけです。抗ヘルペス剤と漢方煎じ薬で治ります。
活性化誘導性T細胞死とは、慢性的な刺激によって必要がなくなった死ぬはずのリンパ球が生き残り、病気を引き起こすといわれるのは実はヘルペスだけは永久に殺し入れないので潜伏感染で細胞の核内に隠れるので殺し切れないので死ぬはずのリンパ球が生き残るのはヘルペスが生きているからです。
免疫反応は時に誤った方向に作用することがあるといわれますがウソです。免疫反応は時に誤った方向に作用することがあるといわれますが病原体を殺し人体を守る免疫系は絶対に生命に遺体して大して危害を及ぼすことは無いのです。
アレルギーは、免疫系が環境抗原である化学物質であるアレルゲンに反応してIgE抗体(もともと寄生虫感染に対処するための抗体)を産生することによって生じるのです。アレルギーやアトピーについてはここを読んでください。
アトピー体質の人は、世界を席巻している化学物質であるアレルゲンを認識しやすい認識しやすい多様性のあるMHC(主要組織抗原複合体遺伝子蛋白質)に化学物質とペプチドに対して結合したアレルギー抗原に対してIgE抗体を作りやすい「遺伝的体質 」を受け継いでいるからです。これらの化学物質は病原体ではないので漢方で免疫を高めることによって免疫寛容で共存できるので治ります。しかし近頃は昔と比べて爆発的に増えているので一つ一つのアレルゲンに対し数多くのアレルゲンに対して免疫寛容になって言うのに時間がかかるようになってきました。アレルギーに対しては製薬メーカーや化学物質製造会社が責任を取るべきですが誰も告発しません。残念です。
この世にあり得ない偽り100%の自己免疫疾患を起こさないためのT細胞分化におけるチェックポイントのウソつき論文の説明。免疫チェックポイントとは、免疫系の調節因子のことです。これらの免疫系の調節因子は、免疫系のT細胞が無差別に自己の細胞を攻撃するのを防ぐための自己寛容にとって重要です。ただし、一部の癌は、免疫チェックポイントの標的を刺激することで攻撃から身を守ることができます。この文章の批判は次回にします。乞うご期待!!!
はじめに
T細胞は骨髄の造血幹細胞に由来し、胸腺での分化の過程で自己と非自己を識別し抗原特異的に活性化する獲得免疫系に固有の能力を獲得する。胸腺は大きく皮質と髄質から構成され、皮質には多くの未熟な胸腺細胞(T細胞系列の胸腺内細胞)が、髄質にはより成熟した胸腺細胞が存在する。これらの胸腺細胞は胸腺内を次々に移動しながら、それぞれの微小環境に特有の上皮細胞や樹状細胞と相互作用する。その過程で、多様な抗原受容遺伝子を発現する細胞集団がひとまず作り出された後、生体にとって有用で、かつ自己に寛容な細胞だけが選択され生存することを許可されるという多段階のチェックポイントを通過する。本稿では、成熟T細胞集団が完成するまでの胸腺内のチェック機構について解説する。
1. 胸腺皮質におけるT前駆細胞の初期分化
骨髄で産生されたT前駆細胞は、皮質髄質境界領域に豊富に存在する血管を介して胸腺へと移入する。これには胸腺の血管内皮細胞が発現する接着因子(P-selectin)、上皮細胞が発現するケモカイン(CCL21、CCL25)の関与が知られている。移入後の前駆細胞は胸腺皮質上皮細胞が産生するIL-7 とDelta-like 4 (Dll4)の刺激によって増殖するとともにT細胞系列への分化を開始する。このとき、RAG(recombination activating gene)の働きによってTCRβ遺伝子のVDJ領域に遺伝子再構成が生じ、個々の細胞が異なるTCRβ鎖遺伝子を保有するようになる。遺伝子再構成の結果、適切なコドンフレームをもち全長TCRβ鎖を発現できた胸腺細胞では、TCRβ鎖と代替α鎖(pre-TCRα: pTα)が会合してpre-TCRが細胞膜上に発現される。pre-TCRの細胞表面への発現は、リガンド刺激なしにシグナルを誘起し、対立遺伝子座の再構成を阻害する。また、pre-TCRシグナルによって共刺激分子であるCD4やCD8の発現およびTCRα遺伝子VJ領域の遺伝子再構成が誘導され、αβTCRを細胞表面に発現するCD4+CD8+(double positive: DP)胸腺細胞への分化が進行する。pre-TCR依存的に分化が誘導されるこのチェックポイントはβ選択と呼ばれる。
2. 胸腺皮質における正の選択
TCR遺伝子の再構成によって作り出されたTCRを発現するDP胸腺細胞集団の中には、自己MHCを認識できず使い物にならない細胞や、自己成分に対して強い反応を示す有害な細胞が含まれる。そのため、これらの細胞を排除して有用な細胞だけを選択的に分化させる機構が存在する。DP胸腺細胞はまず、自身が発現するTCRと胸腺皮質上皮細胞(cortical thymic epithelial cell: cTEC)に提示された自己ペプチド-MHC複合体(self-peptide-MHC complex: spMHC)との相互作用を介して、リガンドに対して低い親和性をもった細胞のみが生存および分化のシグナルを受ける。このチェックポイントを正の選択(positive selection)と呼ぶ。一方、親和性が低すぎたり高すぎる細胞は死滅する。
cTECには、正の選択を誘導する自己ペプチドを産生するための特殊な抗原プロセシング機構が存在することが近年明らかになってきている。プロテアソームは細胞質内タンパク質を分解しMHCクラスI会合ペプチドを産生するが、β5t構成鎖を含む胸腺プロテアソームはcTEC特異的に発現され、CD8+T細胞のレパトア形成に必要なペプチドの産生を担う。胸腺プロテアソーム特有のプロテアーゼ活性がどのような自己ペプチドを産生し、効率的に正の選択を誘導するのかは分かっていない。一方、cTECのリソソームに豊富に存在するcathepsin Lやthymus-specific serin protease(TSSP)はMHCクラスII会合ペプチドを産生することで、CD4+T細胞の正の選択に関与する。また、通常はストレスや飢餓状態にさらされた細胞で生じるオートファジー(autophagy)が、cTECの多くでは恒常的に活性化されており、内因性抗原がオートファゴソームを介してリソソームに運搬されることが知られている。
3. 胸腺髄質における負の選択
胸腺皮質で正の選択を受けて生存したDP胸腺細胞はCD4+CD8-あるいはCD4-CD8+ (single positive: SP)胸腺細胞へと分化誘導されるとともにCCR7を発現し、そのリガンドであるCCL19およびCCL21が豊富に発現される胸腺髄質へと移動する。髄質では、胸腺髄質上皮細胞(mTEC)や樹状細胞に提示されたspMHCに対して高い親和性をもつ細胞がアポトーシスにより排除される。このチェックポイントを負の選択(negative selection)と呼び、中枢性自己寛容の確立に必須である。mTECではゲノム上のすべての遺伝子を低レベルで発現する無差別遺伝子発現(promiscuous gene expression)が恒常的に生じているために、胸腺外のさまざまな臓器に特異的に発現される抗原(tissue-specific antigen: TSA)に対しても自己寛容を確立させることができる。Aire(autoimmune regulator)は無差別遺伝子発現を制御する核内因子であり、その欠損は自己免疫疾患を引き起こす。また、胸腺髄質は制御性T細胞(Treg)の分化の場としても自己寛容に貢献する。spMHCとの親和性が比較的強く、かつ負の選択の閾値を下回る細胞がTregに分化すると考えられている。Tregの分化には、spMHCを認識することによるTCRシグナルに加えて、CD28やOX40、GITRなどのTNF受容体スーパーファミリーを介した共刺激、TGF-βやIL-2などのサイトカインが必要とされる。以上記してきた多段階のチェックポイントを経て髄質で成熟した胸腺細胞は、スフィンゴシン1リン酸受容体1(sphingosine 1-phosphate receptor 1: S1P1)を発現し、血中のリガンド(S1P)に誘引されて胸腺から移出する。
4. 正および負の選択を起こすTCRシグナル
spMHCを認識することで惹起されるTCRシグナルは、その親和性の違いによって、生と死という真逆の運命を胸腺細胞にもたらす。TCRとspMHCとの親和性は結合時間に反映され、親和性が高いほど解離しにくい。この違いは下流で活性化されるMAPK経路の違いとして現れる。正の選択には、低親和性ペプチドの結合によってもたらされるLATの部分的なリン酸化とそれに続くRasを介した弱く持続的なERKの活性化が関与する。一方、高親和性ペプチドによる負の選択ではLATは完全にリン酸化され、Grb2やSOSを介したJNKや p38キナーゼの活性化を引き起こす。様々な強度のTCRシグナルが正の選択と負の選択という対極の結果に帰結する機構として、最近、中程度の親和性をもったペプチドで惹起されるTCRシグナルの強度を特異的に減弱させることで正の選択を促すThemis(thymocyte-expressed molecule involved in selection)の関与が示唆されている。
おわりに
TCR遺伝子の再構成によって作られた初期の胸腺細胞集団から、生体に有用な細胞だけが胸腺内で選択され成熟T細胞となり全身のリンパ組織へ供給される。胸腺細胞の選択はspMHCとの相互作用によってもたらされるTCRシグナルに依存する。最近の研究から胸腺上皮細胞が独自の機構によって胸腺微小環境固有の自己ペプチドを作りだすことが明らかとなってきた。今後、胸腺細胞の選択を引き起こす自己ペプチドの実体が解明されることで、T細胞分化におけるチェックポイントの実像がよりくっきりと見えてくるのではないだろうか。
胸腺細胞は分化の進行に伴って、CD4およびCD8の発現を変化させる。幼若な胸腺細胞はCD4およびCD8を両方とも発現せず(double negative: DN)、DN細胞はさらにCD25とCD44の発現によってDN1からDN4に分類される。DN2からDN3にかけてTCRβ遺伝子の再構成が生じ、TCRβとpTα(pre-TCRα)からなるpre-TCRが細胞表面に発現されることでβ選択が誘導される。このシグナルによってTCRβ遺伝子の再構成は停止され、CD4およびCD8の両方を発現したDP(double positive)細胞への分化およびTCRα遺伝子の再構成が誘導される。TCRαとTCRβから構成されたTCRとspMHCとの弱い相互作用によりDP胸腺細胞は正の選択を受けてTCRα遺伝子の遺伝子再構成を停止するとともに、CD4あるいはCD8のどちらかを発現するSP(single positive)細胞へと分化する。
自己ペプチド-MHC複合体(self-peptide-MHC complex: spMHC)との親和性に依存した胸腺細胞の選択
TCRとspMHCの親和性は、胸腺細胞の運命を決定づける。親和性が全くないか、あるいは非常に低い細胞は生存シグナルを受けられずに死滅する(無視による細胞死、death by neglect)。ほとんどの胸腺細胞はこの運命をたどる。細胞がspMHCに対して親和性の低いTCRを発現している場合には、生存と分化が誘導される(正の選択)。一方、親和性が高い場合にはアポトーシスが誘導されて排除されるが(負の選択)、そのなかでも比較的親和性の低い細胞はTregに分化する(アゴニスト選択)。
T細胞の分化と選択を司る胸腺内微小環境
皮質髄質境界領域の血管から移入したT前駆細胞は、皮質においてDN細胞からDP細胞へと分化する。cTECに発現されるspMHCとの親和性によってDP細胞の生死が決定される。cTECに特異的に存在する胸腺プロテアソームは、細胞質内の自己タンパク質を分解してMHCクラスIに提示される自己ペプチドを産生し、CD8+T細胞の正の選択に不可欠な役割を果たす。同様に、自己タンパク質がオートファジーを介してリソソームへ運ばれると、cathepsin LやTSSPなどのプロテアーゼによって分解され、その結果産生された自己ペプチドはMHCクラスIIに会合してCD4+T細胞の正の選択を誘導する。正の選択の際に惹起されたTCRシグナルによって、SP細胞への分化とともにCCR7の発現が誘導され、胸腺細胞はmTECが産生するリガンドケモカイン(CCL19およびCCL21)に誘引されて髄質へ移動する。mTECには核内因子であるAireが発現されており、TSAの発現を担う。TSA由来の自己ペプチドはmTEC上のMHCに直接提示されるか、あるいはDCに取り込まれたのちに提示される。このspMHCに高い親和性を持つSP胸腺細胞は負の選択により排除されるか、Tregに分化することで中枢性自己寛容が確立される。成熟したT細胞は走遊因子S1P に対する受容体S1P1を発現し、S1Pが豊富に存在する血流中に放出される。

{kind=link}
{kind=link}
{kind=link}