プロスタグランジンはヘルペスと共存するための生理活性脂質である。決して薬理活性物質でもなく、免疫活性物質でもありません。あくまでも生理的な働きを活性化する脂質であります。アスピリンとプロスタグランジンの関係について。特に、プロスタグランジンE2(PGE2)とプロスタグランジンE3(PGE3)の生理活性脂質としての働きを明らかにしましょう。
私は製薬メーカーが作った薬は全て免疫を抑えてしまうので病気を治すことができないと常々考えていました。例えば、ヘルペスとの戦いで痛みが出たときに用いられるアスピリンも使ってはならないと考えていました。なぜならば、ヘルペスによる疼痛は免疫を抑えるから使用してはならないと深く考えずに決めつけていました。言うまでもなく、病原体の中で最も特異的かつ人類最後に残った病気を作り続けるウイルスであるのはヘルペスウイルスであります。ヘルペスとの戦いでアスピリンで疼痛を一時的に消し去ってもヘルペスを殺すことはできないのみならず、ヘルペスは人間の免疫機構から上手に逃げてしまうので世界中の全ての人は多かれ少なかれ一生ヘルペスが感染した状態(潜伏感染)が続くのです。
ところが、アスピリンだけを疼痛軽減のために使ってきたリウマチなどの自己免疫疾患の患者さんとの出会いが何人かありました。リウマチなどの疼痛に対してアスピリンだけを使ってきたそのような患者さんのリンパ球はまったく減少していないのみならず、白血球に対してもアスピリンの影響がまったく皆無であることに気が付きました。リウマチの痛みの原因もヘルペスであるので、抗ヘルペス剤だけが免疫を抑えずして痛みをとる唯一の方法だと考えて癌以外の病気をすべて治せる松本医学を確立したと確信していたのですが、抗ヘルペス剤だけでは十分痛みを取れないときには漢方薬や鍼灸以外にアスピリンを同時併用する価値があるのではないかと考え始めました。
免疫学の専門書物は誰よりも良く読み勉強してきたのですが、アスピリンと免疫についてはどの本に一行も書かれていなかったのです。そこでアスピリンとは何かを初めて本格的に勉強し始めました。アスピリンはプロスタグランジンを産生するCOX-1を抑制する鎮痛剤であることは知っていたのですが、アスピリンと免疫の関わりのみならずCOX-1がどんな酵素であり、どのようにまたどのようなプロスタグランジンの産生を抑制しているのかを完全に理解する必要を感じ、長期間にわたって勉強した成果を皆さんに少しずつお知らせしたいと思います。この論文は未だ完全には仕上がってはいなかったので途中経過を出すことを躊躇したのですが、結論は私の頭の中では出来上がっており、かつ患者さんの要望もあり未完成ながらも例のごとく難しいですが完璧主義者である私に最後まで付き合って下さい。乞うご期待!!
プロスタグランジンは生理活性物質であり、免疫反応をTh-2の世界に進ませ、最後は免疫寛容をヘルペスと共存するために起こす。自然後天的免疫寛容が進化したのは、病原体のヘルペスと共存するためである。ヘルペスは、ワクチンが作れないし、終生免疫も生まれないし、メモリーT細胞も生まれないし、しかも殺しきれないし、人体の高等免疫から回避するテクニックも持っており、一旦、細胞の中に感染してしまうと免疫が正常である限り潜伏感染で隠れ、免疫が弱くなってきた時に増殖し出します。このような類まれなる大天才のヘルペスウイルスに対して人類は免疫で対応することをやめました。それが人類が唯一、病原体に対して免疫寛容を起こすという策を取ったのです。しかしながら、病原体であることは確かなので、生理学的なプロスタグランジンも高等免疫に協力して免疫寛容を起こすシステムを作ったのです。
プロスタグランジン、英語でProsta-glandin、略してPGは,脂質細胞膜を持っている全ての細胞、とりわけ免疫細胞の大食細胞や樹状細胞から産生されるアラキドン酸代謝産物であり、戦っても無駄である殺しきれないヘルペスに対して免疫寛容を起こす以外に、対応できないために進化したのです。なぜならば、NK細胞やキラーT細胞が、ヘルペスが感染した細胞を殺してしまうと、細胞の機能がなくなるだけで、残ったヘルペスウイルスは人間が死ぬまで人間の免疫を回避して生き続けるからです。そのために、ヘルペスに対峙する最高の方法は、生理学的な問題として対処する以外にないのです。その結果、プロスタグランジンの働きは、免疫のサイトカインではなくて、生体の生理的恒常性(ホメオスタシス)を維持し、特異的な病原ウイルスであるヘルペスと共存するための生理活性物質となりました。
それでは、プロスタグランジン(PG)はどのようにして作られるのでしょうか?PGの代表といえるプロスタグランジンE2 (PGE2)について説明しましょう。PGE2は、まず細胞膜の主成分であるリン脂質からホスホリパーゼA2(Phospho-lipaseA2略してPLA2)によってアラキドン酸(Arachidonicacid)が作られ、さらにシクロオキシゲナーゼ(Cyclooxygenase略してCOX)によってPGG2、さらにPGH2が作られます。さらにPGE合成酵素により、PGH2からPGE2が作られます。最後にプロスタグランジンE合成酵素(PGES)によってPGE2が産生されます。免疫系においては主にマクロファージ,樹状細胞のような抗原提示細胞から産生されます。PGE2はヘルペスウイルスの戦いを穏便に済ませるために様々な免疫担当細胞の活性化をコントロールしているのです。PGE2の主な作用機序は、アデニル酸シクラーゼを介した細胞内cAMPの上昇によるものであり、cAMPは炎症性細胞からの化学伝達物質の遊離を抑制し、ステロイド剤には無い抗炎症作用(好中球性炎症)を示すのです。炎症に関わるサイトカインの産生、ケモカインの産生を抑制し、さらにはサイトカイン受容体の発現抑制など、広範な作用を示すのはヘルペスとの戦いを出来る限り早く終結させようとするからです。また、PGE2の作用は、殺しの免疫反応を助けるTh1タイプから、排除と共存の免疫反応を助けるTh2タイプに変えることであります。ちょうど抗体が殺しのIgGから排除と共存のIgEに抗体の種類を変えることを抗体のクラススイッチというように、ヘルパーT細胞が、Th1からTh2に変わる変化を、私はヘルパーT細胞のクラススイッチと名付けています。言い換えると、抗体はリンパ液や血液などの液体に流れている液性免疫ですから、抗体のクラススイッチと言うのは液性免疫のクラススイッチといい、ヘルパーT細胞は細胞でありますから、細胞性免疫のクラススイッチと言い換えたほうがはるかに理解しやすいのです。
ちなみに現代の免疫学の細胞性免疫と液性免疫の使い方の間違いもついでに指摘しておきましょう。現代免疫学は、Th1の働きを液性免疫といい、Th2細胞の働きを細胞性免疫と名付けていますが、論理的には完全に間違っており意味がありません。さらに免疫学の本にはTh1とTh2のバランスがTh2に傾いた時にアレルギーが起こると述べていますが、これも言うまでもなく大間違いですね。アッハッハ!現代医学は、原理的に間違いが多いので臨床においても免疫を抑えるという病気作りに専念せざるを得ないのです。残念ですね!アッハッハ!cAMPについては後で詳しく書きます。
ホスホリパーゼとは何でしょうか?
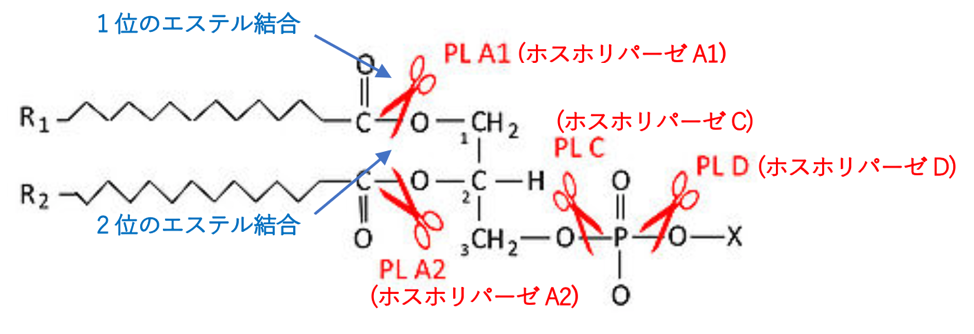
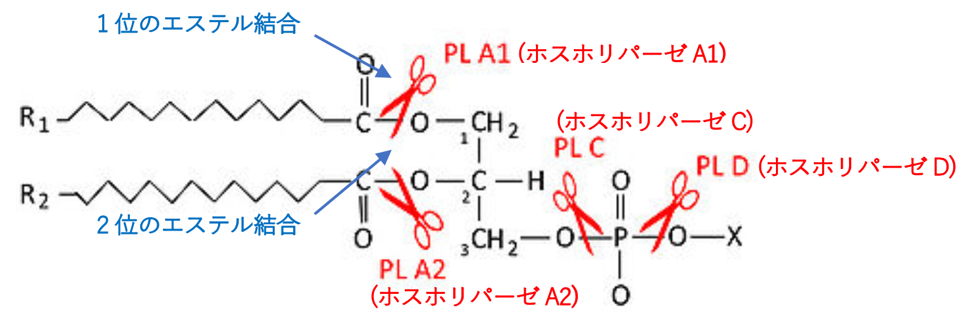
ホスホリパーゼ(phospholipase)は、リン脂質を脂肪酸とその他の親油性物質に加水分解する酵素であります。触媒する反応の種類によりホスホリパーゼA、ホスホリパーゼB、ホスホリパーゼC、ホスホリパーゼDの4種があり、さらにホスホリパーゼAにはホスホリパーゼA1とホスホリパーゼA2の2つがあるので合計5種類あります。ホスホリパーゼA2(PhospholipaseA2)がプロスタグランジンE2を産生するのです。下の絵に、5種類のホスホリパーゼがリン脂質を切断する(加水分解する)部位を示します。ホスホリパーゼA2がアラキドン酸を生成します。
アラキドン酸とは一体なんでしょうか?
アラキドン酸は、細胞膜中のホスファチジルイノシトールのようなリン脂質として存在し、特に脳に多く含まれています。アラキドン酸は、人間が作れない必須脂肪酸であるリノール酸から作られますが、食事からも摂取できます。このアラキドン酸は、ホスホリパーゼA2によってリン脂質から遊離し、ここからエイコサノイドと呼ばれるプロスタグランジンやトロンボキサンやロイコトリエンなどの生理活性物質が作られ、また細胞間のシグナル伝達におけるセカンドメッセンジャーとしても働きます。アラキドン酸からこれらの生理活性物質が作られる生合成過程は、アラキドン酸カスケードと呼ばれます。ホスホリパーゼB(PhospholipaseB)は、ホスホリパーゼA1とホスホリパーゼA2の切断を同時にやり、リゾ(ライソ)ホスホリパーゼ(Lysophospholipase)としても知られています。ホスホリパーゼC(PhospholipaseC)は、リン酸ジエステル結合(グリセロールとの間)を切断し、ジアシルグリセロールとリン酸基を有する頭部を生成します。ホスホリパーゼC(PhospholipaseC)はセカンドメッセンジャーであるジアシルグリセロール(diacylglycerol略してDG、DAG)とイノシトールトリスリン酸(Inositoltrisphosphate略してIP3)を放出し、シグナル伝達において重要な役割を果たしています。ホスホリパーゼD(PhospholipaseD)は、リン酸エステル結合を切断し、ホスファチジン酸(phosphatidicacid)とアルコールを生成します。C型とD型は、実はホスホジエステラーゼと呼ばれることもあります。ホスホジエステラーゼ(Phosphodiesterase略してPDE)とは、広義にはリン酸ジエステル結合(1分子のリン酸が2個の水酸基と結合した構造)の一方の結合を加水分解する酵素です。
リパーゼとエステラーゼの違いはなんでしょうか?リパーゼ(lipase)は、脂質のエステル結合だけを加水分解する酵素群です。一方、エステラーゼ(esterase)は、エステルを水との化学反応で酸とアルコールに分解する加水分解酵素であり、脂質のみならず様々な基質特異性やタンパク質構造を持っている物質や、様々な生理学的機能に応じて、広い範囲の様々なエステラーゼが存在します。上に述べたように、ホスホリパーゼCやホスホリパーゼDのような酵素がリン脂質を分解するエステラーゼの一つとなります。
ホスホリパーゼA2は、完全なレシチン分子に対して作用し、2位の脂肪酸エステル結合を加水分解します。下図のブルーの矢印で示した2位のエステル結合を確認してください。生成物は脂肪酸とリゾ(ライソ)レシチン(lysolecithin)となります。ちなみに、ヘビ毒、ハチ毒(Beevenom)の主成分で哺乳類では膵液中に存在し消化酵素の役割を果たすのがホスホリパーゼA2であります。
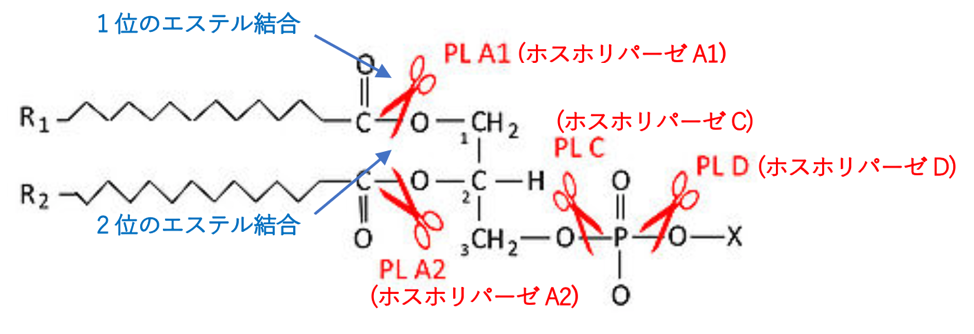
【一般的なリン脂質を加水分解する酵素ホスホリパーゼの種類と、それぞれの酵素が切断する(加水分解する)部位】
の構造式.png)
【リン脂質の1種であるホスファチジルコリン(レシチン)の構造式】
最初に何がアラキドン酸を作らせるのか?
アラキドン酸(Arachidonicacid)は、不飽和脂肪酸のひとつであり、4つのcis二重結合を含む20個の炭素鎖からなるカルボン酸で、ω-6脂肪酸に分類されます。ω-6のωとは何でしょうか?脂肪酸の最後の炭素をωといいます。それではω-6とは何でしょう?最後の炭素から数えて6番目の炭素という意味です。専門的な数値表現でを使えば、20:4(n-6)または20:4(Δ5,8,11,14)で一目瞭然ですが、この表現の意味を説明しましょう。下にアラキドン酸の構造式を図示します。n-6はω-6と同じ意味です。1番左のCOOHはカルボキシル基で、下の青の番号の1〜20までは炭素の番号なのです。しかし、2〜20まではCが描かれていませんが、高校の生物で学ばれたように、Cを書くと面倒な上に煩雑になりますから、書かないのは常識なのです。一方、上の紫の1〜20は一番最後から数えたCの番号です。20:4(n-6)の意味は、最初の20は、アラキドン酸の炭素数が20個あり、4は二重結合が4個あるという意味です。次の(n-6)のnは最後から数えて6番目と7番目二重結合があるという意味です。20:4(Δ5,8,11,14)は別の表示法であり、20は、炭素数が20で、4つの二重結合がありますが、(Δ5,8,11,14)は、(Δ5,Δ8,Δ11,Δ14)を縮めた表記であり、Δは二重結合が存在するという印で、Δ5は5~6に二重結合があり、Δ8は8~9に二重結合があり、Δ11は11~12に二重結合があり、Δ14は14~15に二重結合があることを示します。この表示は生化学を勉強した人にとっては、アラキドン酸を示す一目瞭然の表記法となるのです。
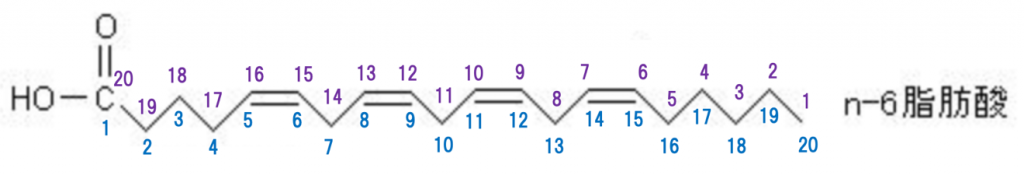
cis(シス)二重結合とは何でしょうか?
ついでにcis(シス)に対してtrans(トランス)という対立語がありますから、それも説明しましょう。
有機化合物における立体異性体の一つであるのが幾何異性体(geometricalisomer)でありますが、実は、正式には、シス-トランス異性体(cis-transisomer)と呼ぶべきなのです。異性体(isomer)とは、同じ数、同じ種類の原子を持っているので同一分子式(同一化学式)となるのですが、構造が異なる物理的、化学的性質の違う同じ物質と言ってもいいのです。幾何異性体(シス-トランス異性体)と呼ばれる異性体には3種類あります。1つ目が、今説明しようとしている有機化合物の二重結合への置換によるシス-トランス異性体。2つ目が、環状化合物の環(リング状)への置換によるシス-トランス異性体。3つ目が錯体配位子の位置の違いによる異性体の3つがあります。
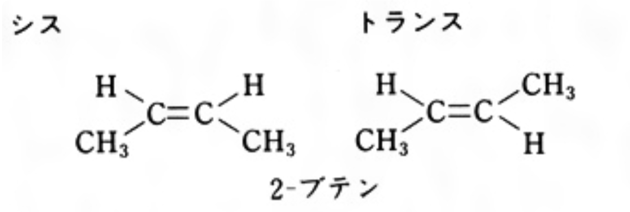
左 の2-ブテンという化学物質を例に二重結合のシス-トランス異性について説明しましょう。まず炭素の二重結合に2つずつのHとCH3の異なった基が結合する場合を例に取りましょう。二重結合を軸として同じ側にHだけとか、CH3だけの2個の置換基がある場合をシス、HどうしCH3どうしが反対側にある場合をトランスと呼ぶのです。上のシスとトランスの図を見れば、簡単に理解できるでしょう。
アラキドン酸とは何でしょうか?
アラキドン酸は、4つのcis二重結合を有する20個の炭素鎖からなる脂肪酸である。メチル末端(ωまたはn)から数えて最初の二重結合が6番目と7番目の炭素の間に位置するため、ω-6(n-6)多価不飽和脂肪酸に含まれ、20:4ω-6と記載されます。アラキドン酸は、主に細胞膜のリン脂質のsn-2位にエステル化されて存在します。リン脂質は、大きく分けてグリセリンを骨格とするグリセロリン脂質と、スフィンゴシンを骨格とするスフィンゴリン脂質の2つが存在します。主にグリセロリン脂質にコリンが結合したホスファチジルコリン(レシチン)に含まれますが、ホスファチジルイノシトールなど他のグリセロリン脂質にも含まれます。遊離アラキドン酸の大半は細胞膜のリン脂質に再度取り込まれます。遊離アラキドン酸はプロスタノイドやロイコトリエンなど多様な生理活性脂質に変換され、摂食、睡眠・覚醒、脳血流など生理的な脳機能を維持する他に、ヘルペスウイルス感染の際の発熱や内分泌応答、疼痛、てんかん、脳虚血、ストレス、神経・精神疾患など様々な病態にも関わっているのです。
細胞膜のリン脂質のsn-2位とは何でしょうか?
snの英語は、stereo-specificnumberingであり、訳すと立体特異的番号付けとなります。主に生化学において脂質を含むグリセロール誘導体の立体配置を記述するための方法です。それではグリセロールとは何でしょうか?グリセロールは皆さんがよくご存じのグリセリンであり、3価のアルコールであり下に4つの構造図と分子式を掲載しました。さらに数多くある異性体を2つ掲載しておきました。3つのOH基があるので、グリセリンは3価のアルコールなのです。ちなみにアルコールを炭水化物と考えておられる人が多いのですが、アルコールは脂肪であることを確認しておいてください。このようにグリセリンは異性体があるので、脂肪酸とエステル結合を行う時に、新たにできた化合物を正確に分別するために生まれたのがsn記号であります。つまり、グリセロールの3つの炭素原子にあらかじめ立体特異的な番号を付けておくことで、立体異性を区別して表現することができるのです。フィッシャーの投影図というのは、私が今まで書いた構造式は全てフィッシャーの投影図であることを知っておいてください。実際、私たちが見る化学物質の構造式の99%はフィッシャーの投影図で書かれています。下の絵図のように1番左のフィッシャー式で中央の炭素原子を2位(sn-2)とし、この2位の炭素に結合する水酸基を左に置いたとき、上の炭素を1位(sn-1)、下の炭素を3位(sn-3)とします。この方法で化合物を命名する場合には、glycerolの直前にsn-接頭辞(stereo-specificallynumberedの意)をつけて明示します。
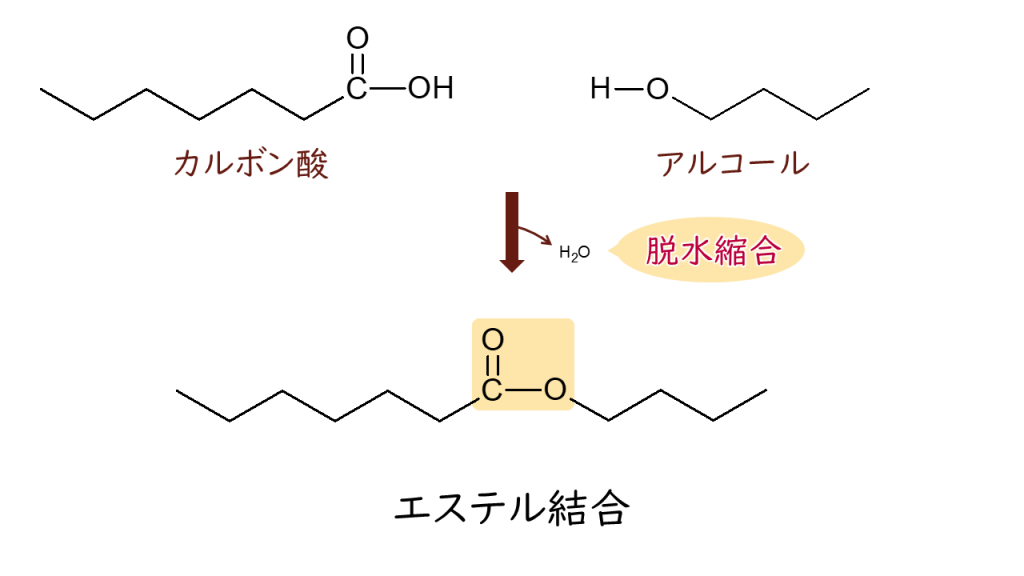
エ ステル結合とは何でしょうか?
左の図のカルボン酸からOHが取れて、アルコールのHが取れて脱水縮合反応が起こり、(-COO-)というエステル結合が起こります。
このように、脂肪酸のカルボン酸とアルコールとの結合をエステル結合といいます。カルボン酸はカルボキシ基(-COOH)をもつ有機化合物であり、アルコールは水酸基(-OH)をもつ有機化合物で、これらの官能基が反応すると、COOHのOHが取れ、アルコールの水酸基のOHからHが取れ、取れたOHとHが結びつくと、水(H2O)となり流れ去り(脱水し)、脂肪酸とアルコールの2つ分子が縮んで合わさるので脱水縮合反応と呼びます。その結果、エステル結合(-COO-)が形成されるのです。脂肪酸はカルボン酸の1つですので、脂質では脂肪酸とアルコールとのエステル結合がよく見られます。
中性脂肪とはなんでしょうか?
グリセロール(グリセリン)と脂肪酸がエステル結合した単純な構造の脂質を、中性脂肪(neutralfat)や、中性脂質(neutrallipid)といい、脂肪酸のグリセリンエステルを指します。脂肪酸のグリセリンエステルは、グリセリン脂肪酸エステルともいいます。モノグリセリド、ジグリセリド、トリグリセリドの3種類が存在しますが、血液検査で既に知っておられると思いますが、血液中に含まれる中性脂肪のほとんどはトリグリセリドなので、中性脂肪はトリグリセリドと考えておられる人が多いのです。トリグリセリドは、英語でTriglycerideと書き、略してTGや、TAGや、Trigという略語で記されます。脂肪酸とグリセリンが結びついて中性を示すので、中性脂肪というのです。中性脂肪は動物の脂肪や植物の油の主成分です。グリセロールはアルコールの1つで、1分子の中に水酸基が3つあります。この水酸基のひとつひとつに脂肪酸が結合できるため、最大3分子の脂肪酸がグリセロール1分子の3つの水酸基に結合します。これが先ほど述べたトリグリセリドなのです。例えば、オリーブオイルに多く含まれるトリオレイルグリセロールという中性脂肪は、1分子のグリセロールの3つの水酸基に3分子のオレイン酸という脂肪酸が結合したものです。
こ のように1分子のグリセロールに3分子の脂肪酸が結合した中性脂肪をトリアシルグリセロール(TG)やトリグリセリドといいます。左の絵図でお分かりのように、中性脂肪には他に、1分子のグリセロールに1分子の脂肪酸が結合したモノアシルグリセロール(MG)、1分子のグリセロールに2分子の脂肪酸が結合したジアシルグリセロール(DG)があります。モノ、ジ、トリはそれぞれ1、2、3を表す倍数接頭辞です。
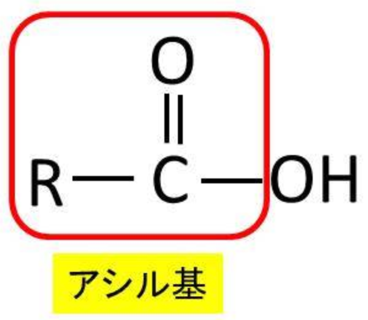
アシル基とは何でしょうか?
上の図で見ると、アシルという言葉がモノアシル、ジアシル、トリアシルの3つともについていますね。アシル基といえば、カルボン酸からOHを抜いた形、すなわち左の図のようにR-CO-という形の基を示します。もちろん-COO-はエステル結合ですが、エステル結合で脂肪酸のカルボン酸COOHとアルコールのOHが反応して、アルコールのHがなくなり、かつカルボン酸のOHがなくなった赤枠で囲んだ残りをアシル基といいます。
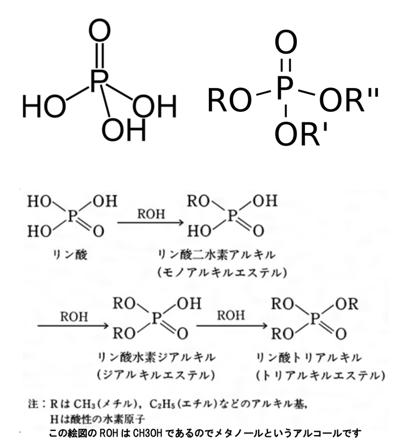
リン 酸エステルとは何でしょうか?
リン酸エステル リン酸エステルは有機リン化合物のうち、リン酸とアルコールが脱水縮合したエステルを示します。左の図でお分かりのように、リン酸の分子式はH3PO4であり、3個の水素の全てまたは一部が有機基で置き換わった構造を持ちます。その置換の数が1、2、3個のものを順にリン酸モノエステル、リン酸ジエステル、リン酸トリエステルと呼び、リン酸エステルも3種類あり、グリセリン脂肪酸エステルと非常によく似ていますね。
リン酸エステルは、リン酸とアルコールの脱水縮合、あるいは、リン酸塩化物とアルコールとを塩基の作用により縮合させることで得られます。また、亜リン酸エステル(P(OR)3など)を酸化するとリン酸エステルに変わります。リン酸ジエステル(ホスホジエステル)は生体においてATPやRNAなどの核酸の部分構造として、生体分子の高次構造や機能に直結する重要な役割を担っていることは皆さんご存じですね。
さて、本文の不飽和脂肪酸の話に戻りましょう。
生体中の脂肪酸の大部分はトリアシルグリセロールとして存在し、エネルギー源として脂肪組織に貯蔵され、これらは電気的に中性であることから中性脂肪と呼ばれることは既に述べました。生体中の脂肪酸の大部分はトリアシルグリセロールとして存在し、エネルギー源として脂肪組織に貯蔵されます。ちなみに、アルコールなどとエステル結合をせずに単独で存在している脂肪酸を遊離脂肪酸(freefattyacid略してFFA)あるいは非エステル結合型脂肪酸(non-esterifiedfattyacid略してNEFA)と呼び、血清中ではアルブミンと結合して溶存しています。ちなみに、常温で融点が低い液状の中性脂肪を油(oil)、融点が高い固体のものは脂(fat)と表現することがありますが、中性脂肪の融点はその構成する脂肪酸の種類によって決まります。例えば、3分子のパルミチン酸をもつトリパルミチンの融点はおよそ66℃、3分子のオレイン酸をもつトリオレインの融点はおよそ5℃になります。なお、脂肪酸の種類は多くあるうえに、1分子のグリセロールに結合する3つの脂肪酸はバラバラでも構わないため、トリアシルグリセロールのバリエーションにはかなり多様性があります。
プロキラルとは何でしょうか?
異性体を説明した時に見慣れないキラリティーという言葉が出ました。キラリティーとは、化学において重ね合わせることのできない鏡像を持つ分子の性質を示します。グリセロールはプロキラルな化合物で、3つの炭素原子のうち中央以外のどちらに置換基が導入されるかによって鏡像異性体を生じます。
鏡像異性体とは何でしょうか?
対掌体ともいいます。対掌というのは、ちょうど右手と左手のように互いに鏡像である1対の立体異性体を持ち、これら2つの異性体は互いにエナンチオマー(enantiomer)、対掌体(対称体は誤り)といい、または鏡像異性体であるといいます。言い換えると、立体構造が互いに実像と鏡像の関係にある一対の異性体を鏡像異性体というのです。不斉炭素原子に結合している原子または原子団の立体的な配置が異なっている2種類の分子は,右手と左手の関係になっていて,互いに重ね合わすことができない.実像と鏡像が光で出現するのでこのような光学異性体は偏光面を回転させる旋光性の方向だけが反対で,他の物理的、化学的性質である旋光度の絶対値、融点、沸点、溶解度、蒸気圧などは全て同じなのです。この関係は不斉炭素原子が存在しない分子不斉の場合でも同じです。しかし、鏡像異性体がほかの光学活性物質と反応する場合には反応速度に差が生じるので、生成物も異なってくることがあります。光学活性物質とは、異性体の違いにより生理活性が異なる物質です。
プロキラリティー(prochirality)とは何でしょうか?
プロキラリティーは、有機化合物の立体化学的性質の概念のひとつであり、ある化合物がプロキラリティーを持つとか、ある化合物がプロキラルである、というのは、その化合物自体はキラリティーを持たないが、しかるべき付加反応、あるいは置換反応を受けることによって、一段階でキラリティーを持つ化合物に変わる、ということを意味します。すなわち、「プロキラリティー」は「キラリティー」の前段階であります。プロキラリティーのもととなる反応点の炭素原子をプロキラル中心炭素と呼びますが、それは2種類に大別できます。一つは、付加反応を受けることでキラル中心になる、「プロキラル面」と呼ばれる平面三角形型炭素、もう一つは、置換反応を受けることでキラル中心になる四面体型炭素です。
再びアラキドン酸について
アラキドン酸は、肉、卵、魚介類などの食品から得られ、細胞内のリン脂質に取り込まれ、様々な生体膜の合成に使用されます。成人では、脳で代謝されるアラキドン酸は血漿から補われ、脳内のアラキドン酸の含有量は一定に保たれています。
アラキドン酸は、必須脂肪酸であるリノール酸か作られ、ナッツなどの種実類や植物油に豊富に含まれています。体内に取り込まれたリノール酸は、段階的な不飽和化および脂肪鎖伸長によって、アラキドン酸やドコサ-テトラエン酸(22:4ω-6)などの脂肪酸に変換されます。ドコサ-テトラエン酸の22:4ω-6の意味はわかりますね。
リノール酸からどのようにしてアラキドン酸が作られるのでしょうか?
まず、リノール酸はΔ6不飽和化酵素(fattyaciddesaturase2略してFADS2)による脱水素化を介して二重結合が付与されてγ-リノレン酸(18:3ω-6)になります。desaturaseというのは、不飽和化酵素の意味です。その後、γ-リノレン酸からΔ6脂肪酸伸長酵素(Δ6elongase)により脂肪酸が伸長されてジホモ-γ-リノレン酸(20:3ω-6)になります。elongaseというのは伸長酵素です。Δ5不飽和化酵素(fattyaciddesaturase1略してFADS1)によりジホモ-γ-リノレン酸からアラキドン酸が産生されます。Δ5不飽和化酵素やΔ6不飽和化酵素の活性は、栄養、喫煙、老化などの要因により変動し、肥満に関与しています。
遊離アラキドン酸はACSLによりアラキドノイルCoA(arachidonoyl-CoA)となって活性化され、アシルトランスフェラーゼ(acyl-transferase)により細胞膜のリン脂質のsn-2位に取り込まれます。sn-2位についてはもうすでに説明しました。リン脂質の産生には二つの生化学的経路が関与します。1つ目はケネディー経路であり、2つ目はランズ回路です。1つ目の経路はグリセロール-3-リン酸アシルトランスフェラーゼ(glycerol-3-phosphateacyl-transferase略してGPAT)がグリセロール-3-リン酸(GP)のsn-1位に脂肪酸をエステル化により付加し、リゾリン脂質の一種であるリゾホスファチジン酸(lyso-phosphatidicacid)を生成します。続いて、リゾホスファチジン酸アシルトランスフェラーゼ(lyso-phosphatidicacidacyl-transferase略してLPAAT)がリゾホスファチジン酸のsn-2位に脂肪酸をエステル化により付加し、ホスファチジン酸(phosphatidicacid)を生成します。ホスファチジン酸はジアシルグリセロール(diacylglycerol略してDAG)に変換され、トリアシルグリセロール(triglyceride)、ホスファチジルコリン(phosphatidyl-choline略してPC)、ホスファチジルエタノラミン(phosphatidyl-ethanolamine略してPE)、ホスファチジルセリン(phosphatidylserine略してPS)が産生される。また、ホスファチジン酸はシチジン二リン酸ジアシルグリセロール(cytidinediphosphate-DAG略してCDP-DAG)にも変換され、ホスファチジルイノシトール(phosphatidylinositol略してPI)、ホスファチジルセリン、ホスファチジルグリセロール(phosphatidylglycerol略してPG)、カルジオリピン(cardiolipin略してCL)が産生されます。
2つ目のランズ回路では、一度生成されたリン脂質をホスホリパーゼA2(phospholipaseA2略してPLA2)によりsn-2位の脂肪酸が遊離し、リゾリン脂質が生成されます。リゾリン脂質アシルトランスフェラーゼは、生成されたリゾリン脂質のsn-2位に脂肪酸をエステル化により付加し、その結果、リゾリン脂質はリン脂質に戻ります。ランズ回路を担うリゾリン脂質アシルトランスフェラーゼ(lyso-phospho-lipidacyltransferase略してLPLAT)には数多くのアイソフォームが存在し、それぞれ基質とするリゾリン脂質や脂肪酸の種類に特異性があります。
細胞膜からどのようにしてアラキドン酸が遊離するのでしょうか?
細胞が成長因子、ホルモン、サイトカインなど様々な細胞外刺激に曝されると遊離アラキドン酸が産生されます。遊離アラキドン酸の産生には、PLA2(ホスホリパーゼA2、phospholipaseA2)による細胞膜からのアラキドン酸の遊離と、アラキドン酸を構造に含むエンドカンナビノイドの代謝による遊離アラキドン酸の産生の2種類のメカニズムが存在します。各経路の機能的意義にも関わる問題であり、今後精査が必要です。細胞膜からの放出後は、遊離アラキドン酸の90%以上は直ちに長鎖アシルCoA合成酵素(ACSL)によってアラキドノイルCoA(arachidonoyl-CoA)となり活性化され、リゾリン脂質アシルトランスフェラーゼにより細胞膜のリン脂質のsn-2位に再エステル化されて再利用されます。リゾリン脂質とは、アシル基を1本有するリン脂質の総称で、リゾリン脂質はグリセロール骨格とスフィンゴシン骨格を有するクラスに大別され,それぞれに結合する極性基とアシル基の種類の組み合わせにより多数の分子種が存在します。リゾリン脂質の物理化学的特徴は,親水性のリン酸基と疎水性のアシル基を有することにあります.
脳内ではアラキドン酸を含むほとんどの多価不飽和脂肪酸は長鎖脂肪酸CoAリガーゼ(long-chain-fatty-acid-CoAsynthase略してACSL)により活性化されエステル化されて、細胞膜のリン脂質になる他に、エネルギー源としてβ酸化により代謝されてアセチルCoAの産生を促します。アラキドン酸の代謝の効率は、細胞膜にある脂肪酸トランスポーターや脂肪酸結合タンパク質(fattyacid-bindingprotein略してFABP)によります。
アスピリンやインドメタシンなどの非ステロイド系抗炎症薬(NSAID、エヌセイドと発音します)が、シクロオキシゲナーゼ(COX)の活性を不可逆的に阻害してその薬理効果を発揮します。プロスタグランジンは古くから炎症性のメディエーター(仲介者)として知られていましたが、その意味づけがなされていませんでした。その意味づけを、今私が明らかにしようとしているのです。生理的あるいは殺しきれないヘルペスを含む腸の常在病原体刺激に応じて産生、分泌され、標的細胞であるTh0をTh1から出来る限り早くTh2に変換するように作用し戦いのない生体のホメオスタシスの維持および免疫寛容をもたらす生理活性脂質なのであります。
プロスタグランジンの産生は,細胞膜リン脂質からホスホリパーゼA2(PLA2)によって切り出されたアラキドン酸が,COXによってPGG2となり、さらにPGH2になり,最後はそれぞれのプロスタノイドに特異的な合成酵素によって,生理的に重要な5種類のプロスタノイド(PGD2、PGE2、PGF2、PGI2、TXA2)が5種類合成され,細胞外に放出されます。これらプロスタノイドの生理活性は標的細胞上の特異的な受容体(Gprotein-coupledreceptor略してGPCR訳してGタンパク質共役受容体)を介して発揮されます。PGD,PGE,PGF,PGI,TXの受容体はそれぞれDP,EP,FP,IP,TPです。プロスタノイドの中でも多彩な生理活性を持つPGEについては4種類の受容体サブタイプが存在しており,EPI,EP2,EP3,EP4に分類されます。これら受容体はすべてG蛋白質共役型(GPCR)で,網膜にある光受容体であるロドプシンタイプ7回細胞膜貫通型受容体構造をしています。共役する(連結している)情報伝達系の種類から,3つのグループに分けられ,EP1,FP,TPはCa2+動員系に,EP2,EP4,DP,IPはアデニル酸シクラーゼの活性化に,EP3はアデニル酸シクラーゼの抑制に共役連結しています。
PGE2(プロスタグランジンE2)の免疫担当細胞に対する作用は、全てヘルペスとの戦いをやめて共存するという合目的な目的を持っています。以下にその証拠を具体的に説明していきましょう。例のごとく難しいのは言うまでもありません。
免疫系とプロスタグランジン
免疫系においても様々なプロスタグランジン(PG)がプロスタノイドの標的細胞上の特異的な受容体(Gprotein-coupledreceptor略してGPCR訳してGタンパク質共役受容体)を介して発揮されるのです。とりわけPGEは免疫担当細胞の中でも特にマクロファージ、樹状細胞のような抗原提示細胞から産生され、抗原提示細胞自身およびその周辺細胞(T、B細胞)に影響を与えています。
PGE2が免疫担当細胞であるT細胞、B細胞、樹状細胞、マクロファージに及ぼす作用について詳しく述べましょう。
まずサイトカイン産生工場であるT細胞に対する作用については、
1)病原体を殺す仕事をするTh1やNK細胞に対して、PGE2は免疫をできる限り戦わせないようにするために、戦いのサイトカインであるIFN-γとIL-2の産生を抑制します。
2)さらにTh0から戦いのTh1への分化抑制をさせるのみならず、化学物質とヘルペスと共存させるTh2への分化を亢進させます。これは、Th2は排除の戦いであり、かつ素早くTh2に変えてTh2にIL-10を出させて、Th0をTreg細胞にできる限り早く分化させ、ヘルペスと化学物質を免疫が戦わないで共存させるようにしてくれます。
3)マクロファージや樹状細胞やNK細胞やTh1などが発現しているIL-12レセプターを減らそうとします。というのは、マクロファージや樹状細胞はオートクリンでもあり、パラクリンでもあるTNFを最初に作って、自分に対してIL-12を作れ作れと命令すると同時に、かつIL-12はNK細胞に対してもこのIL-12を用いて、IFN-γを作れと命令します。ところが、IL-12レセプターをプロスタグランジンE2が作らせなくなると、IL-12はIL-12のレセプターと結びつくことができなくなり、ますます殺しきれない病原体であるヘルペスとの戦いを抑制し、ヘルペスと共存するようにさせるのです。さらに、IL-12はTh1に殺しのTNFやIFN-γやIL-2を作らせるのはご存じでしょう。
IL-12についてまとめておきましょう。IL-12はAPC(抗原提示細胞)の役割をするB細胞および単球系細胞(大食細胞や樹状細胞)から産生され、病原体殺しに関わるT細胞やNK細胞に対して細胞増殖の促進、キラーT細胞の細胞傷害活性を高め、IFN-γ産生を亢進する作用も持っています。
4)CD8+CD4+T細胞のアポトーシスするように誘導します。それはPGE2はとにかくヘルペスと戦わせないようにするためです。胸腺でCD8とCD4の両方のタンパクを発現している未熟なT細胞をダブルポジティブT細胞(CD8+CD4+T細胞)は、分化するとCD8+だけを持つかCD4+だけを持つシングルポジティブなT細胞に分化し、CD8+はキラーT細胞になり、CD4+のT細胞はヘルパーT細胞に成熟するのです。ヘルペスや化学物質との戦いを辞めさせようとすれば、CD8+CD4+T細胞をアポトーシスさせてしまえばいいのですが、PGE2は全てのCD8+CD4+T細胞をアポトーシスをさせるわけではありません。
5)Th2に対してはGATA-3を活性化して、Th2のIL-5産生を増強してアレルギーを起こす好酸球を増やします。
GATA-3とは何でしょう?
GATA-3 遺伝子によってコードされる転写因子です。皆さんご存知の通り、Th2細胞はアレルギーを起こす一連のサイトカインIL-4、IL-5、IL-10、IL-13を産生するサイトカイン工場です。まずIL-4が肥満細胞から作られるとTh0がTh2になり、上にあげたIL-4、IL-5、IL-10、IL-13を用いてBリンパ球をIgG抗体からIgE抗体へとクラススイッチさせて、アレルギー反応を起こさせます。これらのサイトカインはTh2サイトカインとよばれますが、もっと正確には殺しきれない化学物質とヘルペスとの共存のためのサイトカインと言うべきです。IL-4、IL-5、IL-10、IL-13のそれぞれをコードする遺伝子は人を含む哺乳類では同一の染色体に近接して存在し同調して発現します。この分化過程を制御するマスターレギュレーターとしてGATA-3が存在しています。要するに、PGE2は転写因子GATA-3をオンにして戦いのTh1を最後は共存できるTh2の世界に変えるために存在しているのです。
PGE2の働きで最も強い作用は殺しのTh1を増えないようにすることであり、細胞内サイクリックAMP(cAMP)の上昇により、Th1サイトカインであるIL-2とIFN-γの産生を抑制するからです。また、細胞内サイクリックAMP(cAMP)はTh2細胞からのIL-5の産生を増強します。上で述べたGATA-3はTh0からTh2の分化に転写因子として必要であり、生理活性資質であるPGE2はcAMPを上昇させ、GATA-3を増強しTh2への分化を促進する手助けをするのです。一方、Th0からTh1への分化のサイトカインとしてはIL-12が必要ですが、PGE2はT細胞上のIL-12受容体の発現を抑制することは既に述べました。極めて難解であるので、全てを理解し記憶するのは素人には無理ですが、興味のある人はついてきてください。
B細胞に対しては、PGE2はIL-4が存在するとB細胞のIgEの産生を促進します。
1)真上ですでに述べたように、IL-4によって抗体のIgGのIgEへの増強作用がPGE2にあります。
2)未熟B細胞のアポトーシス誘導もかかわっています。これもB細胞が成熟するとまず病原体であるヘルペスと無駄な戦いをするので、成熟Bリンパ球を作ると戦いのIgGがまず作らせないようにさせているのです。
樹状細胞とマクロファージに対するPGE2の作用は、IL-12やTNF-αを産生させないようにします。
1)Th0をTh1に変えるIL-12やTNF-αを産生させないようにするのは、殺しのTh1の世界に変えてもヘルペスは殺しきることができないので、最後は共存のTh2へと変えようするのがPGE2(プロスタグランジンE2 )です。要するにヘルペスはTh1であろうがTh2であろうが最終的には戦わないで共存することだけが最良の戦術となるからです。日本はアメリカと中国のいずれと戦っても負けますから、フィリピンのように両国と共存するしかないのと同じです。
2)IL−10産生の増強です。とにかくIL-10はアレルギーの世界であり、かつTregの免疫寛容の世界であることは皆さんすでにご存じでしょう。一旦、敵をTh1の世界で殺しきれないことがあれば次の目標はTh1をTh2に変え、最後の目的はTh0をTregに変えることによってヘルペスと仲良くすることしかないのです。この意味でもプロスタグランジンE2 (PGE2)は、なんと賢い平和的で生理的な活性脂質でしょうか!!言い換えると、ヘルペスに対する無駄な免疫の戦いをやめて平和的かつ生理的な手法で穏便にヘルペスに対処しようとしているのです。
3)IP-10(CXCL10)産生抑制です。IP-10は、英語で Interferon-γ inducible protein 10です。病原体が大食細胞に感染すると、TNFが作られ、このTNFの命令によってNK細胞に殺しのサイトカインであるIFN-γを作らせます。さらにIFN-γによってTh0がTh1に分化し(これを私は細胞クラススイッチと名付けています)、IFN-γがTh1細胞によってもさんせいされます。このIP-10(Interferon-γ inducible protein 10)は、病原体やIFN-γによって、IFNによって刺激された単球(大食細胞や樹状細胞)や線維芽細胞や内皮細胞によって誘導されるケモカインであるので、CXCケモカイン10(CXCL10)と名づけられました。PGE2は、IP-10(CXCL10)の産生を抑制して、免疫の殺しの世界を避け、病原体との戦いによる炎症が起こらないようにしているのです。
4)MDC(CCL22)発現増強です。MDCは英語でMacrophage Derived Chemokine、日本語で、大食細胞由来ケモカインと訳されます。MDCは大食細胞のみならず、B細胞、単球、NK細胞やCD4を持っているT細胞(ヘルパーT細胞、略してTh細胞)などでも産生されます。MDCはCCL22とも呼ばれるのは、CCケモカインの仲間の一つであるからです。MDCのレセプター(受容体)は、CCR4です。CCR4 は Th2 細胞に極めて選択的に発現しています。単球、樹状細胞やNK細胞が病原体であるヘルペスと戦っている部位では、これらの細胞が大量のケモカインを作っています。ケモカインは、正式名はchemo-attractant cyto-kineであり、縮めて先頭と語尾をとってchemo-kineといい、日本語では化学誘引物質と訳します。何をどこに誘引する化学物質であるかというと、免疫細胞がヘルペスと戦っている部位で、免疫細胞がさらに多くの味方の軍隊を呼び寄せる化学物質なのです。つまりケモカインが存在する部位へ走って集まる(遊走)ようにするサイトカインです。ところがMDC(CCL22)と選択的に強く結びつくレセプターがTh2に発現しているのです。ケモカインによって集まってきた免疫細胞の一つであるTh2のCCR4にMDC(CCL22)のケモカインと結合してしまうと、ケモカインの働きがなくなり、免疫細胞を集めることができなくなって炎症反応が起こらなってしまうのです。元来、Th2はアレルギーを起こすT細胞ですから、殺しの戦いのみならず、アレルギーの戦いも起こらなくなってしまうのです。PGE2は、殺しのTh1の世界から、Th2の世界へ出来る限り早く変えて、化学物質やヘルペスを免疫寛容(共存)させようとするのみならず、アレルギーのある人は症状が出ないようにしているのです。ケモカインやケモカインレセプターについてはこちらを読んでください。
5)はCD86(B7.2)の発現増強です。CD86(B7.2)とはいったい何なのでしょうか?まず共刺激について話を始めましょう。共刺激は英語でco-stimulationといいます。実は、抑制刺激もあるので、co-stimulationという表現は片手落ちな説明であることを知っておいてください。もう一つ新しい言葉が、共抑制というべきco-inhibitionであります。順々に説明していきます。
T細胞は敵がT細胞受容体(TCR)に結合するだけでは、戦いを始めたり、つまり戦いをするように刺激されたり、逆に戦いを終わらせる、つまり戦いの刺激が抑制されたりするのです。ところが、必ずもう一つの非特異的な補助的なAPC(抗原提示細胞)からの手助けが必要なのです。元来、APC(Antigen presenting cell)は、特異的な抗原を認識するTCR(T cell receptor)と結びつくだけで、T細胞は活性化できると思い込まれていましたが、実はそれだけではT細胞には何の変化も起こらないことが発見されました。つまり、APCが持っている刺激や抑制が必要であるということが分かったのです。APCからの非特異的な別の手助けを受けたT細胞は、その手助けによってT細胞が刺激にしろ抑制にしろ活性化され始めるのです。この手助けは受け取る側(T細胞)と手助けを与える側(APC)のそれぞれから異なった分子が結合する必要があるのです。
さらに次の段階は、APCからの手助けを得た後、続いてT細胞は自分自身を刺激(抑制)するという2段階の刺激(抑制)が隠されているのです。ところが現代の免疫学の専門用語はこの2段階の刺激(抑制)を、非常に混乱を招く1つの言葉で説明しようとしています。この2段階の刺激(抑制)を現代の免疫学では英語では一言でco-stimulationといい、日本語では共刺激と訳し、かつ、この共刺激に関わる分子をco-stimulatory moleculeと表現するだけですから、ますます2段階の刺激に関わる分子が何なのかが分かりにくくなってしまうどころか、co-stimulationという概念さえもが不明確になってしまうのです。なぜならば、刺激だけがあるのではなくて、抑制の意味が消されてしまったのです。以前、私は共刺激の共はT細胞もAPCも同時に共に相互に刺激され合うから「共」と苦し紛れに説明したことがありますが、この解釈は正確ではなかったことをお詫びします。
以上に述べた補助刺激(共刺激)というのは2種類あって、1つはAPCがTリンパ球を非特異的補助的に刺激(抑制)するという意味と、APCに刺激(抑制)されたTリンパ球が抗原と結びついたTCRを特異的刺激(抑制)するという2つの意味があるのです。この2つの補助刺激を明確に区別するために、私は新しい言葉を作りました。まずco-stimulation(共刺激)という言葉を失くして、「APCによる非特異的、非抗原刺激」(抗原による特異的な刺激ではない)と言うべきです。APCを入れることによって何が刺激していることがわかると同時に、非特異的刺激という言葉を入れることによって特異的な抗原刺激とは関わりがない補助刺激という意味を示したいのです。しかもさらに共刺激分子(co-stimulatory molecule)という言葉を廃止して、2つの言葉に分けるのです。1つはAPCの能動的に刺激する分子をstimulating molecule(刺激する分子)、T細胞の受動的に刺激される分子をstimulated molecule(刺激される分子)とまず分けるべきなのです。stimulating moleculeは刺激するAPCの分子であり、stimulated moleculeは刺激されるT細胞の分子となります。
さらに実はstimulationという言葉の中に2つの意味が含まれていることが問題です。1つはまさに戦いを始めるという意味で刺激する(stimulating)という意味と、2つ目は戦いをやめさせる(inhibiting)という意味を同時に持っているので混乱が起こるのです。免疫学の揺籃期にはTCR(T cell receptor)に抗原が結びつくとどんどん刺激されて、敵である抗原をやっつけることだけしかT細胞の働きはないと考えられていたのは既に述べました。ところが免疫学が進歩すればするほどT細胞は敵を攻撃するだけのみならず、化学物質やヘルペスのような敵と戦いをやめて共存するための新たなるT細胞の働きが分かりだしたのです。
例えばノーベル賞を授与された抗ガン剤であるオプジーボも、活性化されたキラーT細胞にPD-1というレセプターが生まれるので、ガン細胞やヘルペスウイルスが感染した細胞をキラーT細胞が殺す免疫の力がなくなるので、活性化されたキラーT細胞のPD-1に引っ付くオプジーボという人工抗体を投与すると癌細胞を殺す力が復活するのと同時に、ヘルペスが感染した細胞も殺してしまうので、とんでもない数多くの副作用が出現することは、皆さんご存じでしょう。つまりキラーT細胞は殺し屋専門であるのみならず、敵によっては平和的に共存してくれる優しいT細胞の側面も持っているということを伝えたいのです。オプジーボについては私のこの論文を読んでください。
もっと具体的に説明すると、T細胞のPD-1は癌との戦いをやめさせるというinhibition(抑制力)を持たす分子の存在がわかり、そのたびにガン細胞を攻撃する刺激もなくなってしまうということが分かってオプジーボが誕生したのです。キラーT細胞のTCRで癌細胞を認識してもPD-1ができてしまうと殺しの戦いをやめさせるように、stimulatory moleculeの反対の分子であるinhibitory moleculeもT細胞がもつことがあるとわかったのです。従ってこのようなPD-1分子をinhibitory moleculeと名づけるべきなのです。実はこのような分子のことをまた訳の分からない「チェックポイント(immune checkpoint)」などと名づけたりしているのです。差し詰め日本語に訳せば、免疫阻止ポイントとなるでしょう。わかりやすく言えば免疫阻止ポイントは免疫の殺す働きを阻止するという意味です。
したがってTCRが認識する敵に対して2種類の補助分子によって働きがまったく正反対になることが分かったのです。従来からわかっていた1つ目の刺激性経路は殺すべき敵を見つけて俄然興奮する刺激活性化経路であり、2つ目はT細胞が逆にその殺しの戦いをやめさせるための共存の経路を活性化させるのですが、見方を変えれば刺激活性経路を遮断してしまうことです。ところが、それぞれの経路を活性化させるためにはTCRに抗原が結びつくのみならず、この2つのどちらかの経路、つまり刺激経路と抑制経路を活性化させる補助シグナルが絶対に必要なのです。決して共シグナル(共有するシグナル)が必要ではないのです。
1つ目の戦うために必要な補助シグナルをT細胞に与える分子をstimulatory分子(刺激分子)、あるいはstimulating moleculeと呼び、2つ目の戦いをやめさせる補助シグナル分子をinhibitory分子(抑制分子)、あるいはinhibiting moleculeと呼ぶべきなのです。具体的には、戦いのためのT細胞がもつ刺激される分子(stimulated molecule)はCD28であり、このT細胞と結びつくAPCが持つT細胞を刺激する分子(stimulating molecule)はCD80別名B7-1といいます。一方、戦いを止めて共存するためのT細胞がもつ刺激される分子(stimulated molecule)は同じCD28でありますが、このCD28に結合するAPCがもつ抑制する分子(inhibiting molecule)はCD86別名B7-2であります。
このように刺激分子はT細胞とAPCにあり、刺激する分子と刺激される分子同士が結びついて初めてT細胞が殺すために活性化したり、反対にT細胞が共存のために殺しの活性を停止させる場合の2つがあるのです。このような例はいくつかすでに発見されています。3つの例を示しましょう。T細胞のもつ抑制される分子(inhibited molecule)がCTLA4別名CD152 に対して、APCがもつ抑制する分子(inhibiting molecule)は先ほどのCD86別名B7-2であります。2つ目がT細胞にあるstimulated分子の4-1BB 別名CD137に対して、刺激する方はAPCにあるstimulating分子の4-1BBLであります。最後の3つ目はT細胞にあるstimulated分子であるCD154は別名CD40リガンドまたはCD40Lとも呼ばれ、APCにあるstimulating分子はCD40であります。このCD40L(CD154)は、主に活性化T細胞に発現するタンパク質で、分子のTNFスーパーファミリーのメンバーです。抗原提示細胞上のCD40に結合し、標的細胞の種類に応じて多くの刺激活性効果をもたらします。
CD86(B7‐2)とCD80(B7‐1)は何でしょうか?
さて本文に戻ってCD86について説明しましょう。同時にCD86によく似た分子であるCD80についても説明しましょう。CD86(B7‐2)とCD80(B7‐1)は、抗原提示細胞(APC)に発現する免疫チェックポイント分子(免疫刺激阻止分子)であり、B7ファミリー分子とも呼ばれ、役割の異なった2種類のB7‐2分子、B7‐1分子があります。T細胞の活性化において最も重要な刺激する分子(stimulating molecule)がCD80であり、T細胞の抑制化において最も重要な抑制する分子(inhibiting molecule)であるのがCD86なのです。抗原特異的T細胞に非特異的な補助刺激シグナル、もしくは非特異的な補助抑制シグナルを送る主要分子であります。このような補助刺激、もしくは補助抑制シグナルを送る分子はいくつかあることを知っておいてください。
非特異的なCD80(B7-1)はB細胞に発現し、CD86(B7-2)は単球や樹状細胞などの専門的な抗原提示細胞(antigen presenting cell略してAPC)上に発現し、T細胞上のCD28と結合することは既に述べました。CD28とCD80(B7-1)と結合するとCD28からのシグナルがT細胞の核に伝えられてT細胞は刺激され、T細胞の増殖、T細胞から様々なサイトカインの産生、細胞の生存を促進するなどのアクセル作用を発揮する一方、CD28とCD86(B7-2)と結合するとCD28からのシグナルがT細胞の核に伝えられてT細胞の働きが抑制されるブレーキ作用が発揮されることがわかり始めました。
最後に、なぜco-stimuletionを日本語に翻訳する時に不正確にも共刺激と訳したかご存じですか?coという英語の接頭語は、2種類の意味があります。1つ目が、共同の、共通の、相互の、という意味であり、2つ目が、副とか、補助の、という意味があるのですが、最初の日本語に訳す時に日本の偉い学者が、1つ目の「共」という訳にしてしまったので、「補助」という意味がなくなってしまい、その結果、混乱を招く「共刺激」という言葉が生まれてしまったのです。残念です。アッハッハ!
さて、上の文章は極めて難しすぎるので、視点を変えながら話しましょう。PGE2はあくまでも生理活性脂質であって、敵を殺すとか敵と戦うという免疫のメインストリートには直接関わりがなく、ひたすら殺すことができない化学物質とヘルペスウイルスとの共存を目指す作用を発揮するのに生まれた生理活性物質であります。生理学(Physiology) とは、人体を構成する各要素(細胞、組織、器官)がどのような活動を行っているかを解き明かす学問であり、細胞、組織、器官の3つの要素がどのような機能を持つかを明らかにし、その機序への理解を深めていく学問であり、人体に病原体や異物が入ったときにどのように敵を処理するかについては全く興味がないのです。それどころか化学物質に対しては自然後天的免疫寛容を目指して共存させ、病原体である殺しきれないヘルペスウイルスに対しても、出来る限り戦うことをやめて共存できる敵に対しては、平和的に共存するために進化し続けてきたのが生理的な資質であるPG(prostaglandin)なのです。
それではなぜPGE2は発熱や疼痛を引き起こすのか
PGE2は決して免疫のタンパクでもなく、サイトカインでもなく、IFNでもなく、インターロイキンでもないにもかかわらず、なぜ発熱や疼痛を引き起こすのかについてさらに詳しく説明しましょう。だからこそ、PGやPGE2は免疫関連物質ではないので、免疫学に関わる全ての専門的学術書においては、PGについてはほんのわずかしかページが割かれていないのです。というよりも、医学者である免疫学者はPGについては一瞥も与えていないと言ってもいいぐらいです。ましてやPGと免疫の関係を意味づける書物は1冊も存在しないのです。要するにPGは免疫の関わりにおいて何のために存在しているのか、免疫におけるPGの意義づけを明らかにしたいのです。PG(prostaglandin)の代表としてPGE2を中心に詳しく説明していきましょう。生理学的物質であるPGは、本当は免疫との関わりは持ちたくなさそうですが、仕方なく無駄な戦いだけは辞めましょうという平和主義者の仕事をせざるを得なくなったのです。
免疫性感染性発熱の神経回路とPGE2についてまず説明しましょう。
まず、プロスタグランジンには5種類あり、この5種類をまとめてプロスタノイドといいます。この5種類はPGD(プロスタグランジンD2)、PGE2(プロスタグランジンE2)、PGF2α(プロスタグランジンF2α)、PGI2(プロスタグランジンI2)、TXA2(トロンボキサンA2)です。この5種類が結合する受容体というのは、PGD2に対してはIP、PGE2に対してはEP1,EP2,EP3,EP4の4つがあり、PGF2αにFPがあり、 PGI2に対してはIPがあり、TXA2 に対してはTP ,EP2,EP3の5つがあります。難しいですが、興味のある人は付いて来て下さい。
さて病原体による感染が起こると免疫系が活性化され、サイトカイン類が産生され血中に流れ出ます。これらのサイトカインのTNFやIL-12が脳の血管の内皮細胞へ作用すると、内皮細胞内でシクロオキシゲナーゼ-1(cyclo-oxygenase-1、略してCOX-1)やシクロオキシゲナーゼ-2(cyclo-oxygenase-2、略してCOX-2)などの上に述べた5種類のプロスタグランジンに特異的なプロスタグランジン合成酵素群が産生され、発熱メディエーター(仲介者)であるプロスタグランジンE2(prostaglandin E2、略してPGE2)も特異的な酵素によって作れられます。プロスタグランジンE2は脳実質内へ拡散し、脳の視床下部の視索前野のニューロン(神経細胞)に存在するプロスタグランジンEP3受容体に作用します。EP3受容体は抑制性のGTP結合蛋白質と共役するので、結果的に視索前野のニューロンは抑制されます。このような受容体とGTP結合蛋白質が共役する(協力する)受容体のことをG protein-coupled receptor略してGPCRといい、Gタンパク共役受容体といいます。
Gタンパク共役受容体(GPCR)とは何でしょう?ますます難しくなりますが着いて来て下さい。
プロスタグランジン(プロスタノイド)を産生するために用いられるGタンパクには3種類あり、この3種類をGタンパクのサブユニットといい、Gsタンパク、Giタンパク、Gqタンパクの3種類です。TNFやIL-12などの最初の一次メッセンジャーであるサイトカインが受容体に結びつくと何が起こるのでしょうか?これらの受容体と結びついてGタンパクと協力すると何が起こるのでしょうか?どんな働きをするのでしょう?アデニル酸シクラーゼを活性化することです。アデニル酸シクラーゼは英語でadenylate cyclase略してACと書き、アデニリルシクラーゼやアデニレートサイクレースと呼ぶこともあります。アデニル酸シクラーゼはATPを3′,5′-環状AMP (cAMP) とピロリン酸へ変換する反応を触媒する酵素であります。このような酵素を脱離酵素(リアーゼ)といいます。3′,5′-環状AMP (cAMP) とピロリン酸については後で詳しく書きます。
GPCRのGはグアニンのGですが、GPCRというのは実はグアニンヌクレオチド結合タンパク共役レセプターと訳すべきなのです。GPCRの別名は、Gタンパク質結合受容体やあるいは細胞膜を7回貫通する特徴的な構造から7回膜貫通型受容体と呼ばれることもあります。Gタンパク質共役受容体は、生体に存在する受容体の形式の1つである。様々な機能を持ったGタンパク質共役受容体が見られ、既知のタンパク質の中では最大のスーパーファミリーを形成しています。
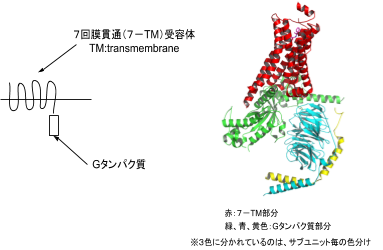
GPCRは受容体とGタンパクが結合した構造をしており、左に2つの絵図を掲載しておきます。左図の左はGPCRのイメージ図です。左図の右は実際のタンパク質結晶のリボン図です。左の左の図は7回膜貫通をしている受容体とGタンパクが結合している状態を示しています。7回膜貫通を英語で、7
trans-membraneといい略して7TMとなります。右の右の図は赤は7TMの受容体の結晶であり、緑、青、黄色はGタンパク質の3つのサブユニットでありGs、Gp、Giにであります。受容体と共役する3つのサブユニットのGタンパクのGs、Gp、Giはすでに触れました。それではGs、Gp、Giの意味について説明しましょう。
まず1つ目のGsタンパク質のsはstimulate(刺激)の略です。このGsタンパク質と共役する受容体にTNFやIL-12などの最初のメッセンジャーなどのサイトカインが細胞膜に結合すると、Gsタンパク質を介してアデニル酸シクラーゼ(adenylate
cyclase略してAC)が刺激(活性化)されます。
アデニル酸シクラーゼが活性化されると、細胞内サイクリックAMP(cAMP:cyclic
AMP)が増加します。細胞内cAMPが増加すると、様々な生理的反応が引き起こされます。受容体にサイトカインのメッセンジャーがやってきたというメッセージを細胞内に伝える役割に注目しcAMPをセカンドメッセンジャーと呼びます。細胞内サイクリックAMPはエネルギー通貨であるATPが酵素であるアデニル酸シクラーゼによってcAMPとピロリン酸に分解されます。cAMPの構造は下の絵図に示しておきます。
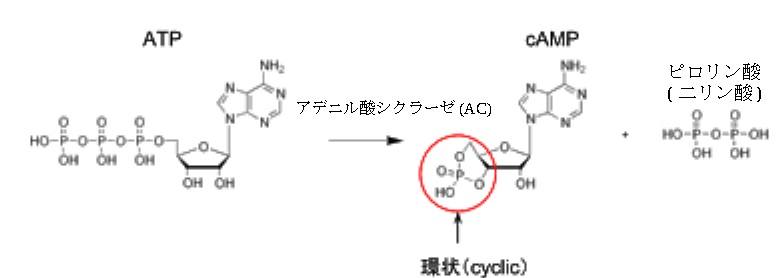
cAMPはアデニル酸シクラーゼにより、ATPから合成されるセカンドメッセンジャーで、Gsタンパク質と共役する受容体(アドレナリンβ受容体やグルカゴン受容体など)は、一次メッセンジャーであります。IL-12やTNFなどが受容体に結合すると、Gsタンパク質を介してアデニル酸シクラーゼを活性化しcAMPを生成します。細胞内でcAMPはプロテインキナーゼAを活性化します。プロテインキナーゼAは英語でprotein kinase Aといい、略してPKAとかAキナーゼとか日本語でcAMP依存性プロテインキナーゼといいます。このPKAが活性化されるとカスケード的に細胞内応答を引き起こすが、細胞によりその応答はさまざまであります。難しいですが着いて来て下さい。
2つめはGiタンパク質です。iはinhibit(抑制)の略で、Giタンパク質と共役する受容体に基質であるcAMPが結合するとGiタンパク質を介してアデニル酸シクラーゼ(AC:adenylate
cyclase)が抑制されます。するとcAMPはホスホジエステラーゼ(PDE)の作用により加水分解されてAMPとなり不活性化されてしまいます。したがってアデニル酸シクラーゼが抑制されると、細胞内サイクリックAMP(cyclic
AMP略してcAMP)が減少します。細胞内cAMPが減少すると、さらに様々な生理的反応が引き起こされるのです。
3つめはGqタンパク質です。qの英語の由来ははっきりしません。Gqタンパク質と共役する受容体に基質であるセロトニンやアドレナリンやオピオイドやヒスタミンやグルタミン酸が結合するとGqタンパク質を介してホスホリパーゼCが活性化されます。ホスホリパーゼCは、細胞膜を原料としてジアシルグリセロール(DG)とイノシトール三リン酸(IP3)を生成します。これらがセカンドメッセンジャーとなって、様々な生理的反応が引き起こされます。以前書いたことがありますが、ホスホリパーゼCがリン脂質のどの
部分を切断するかについての絵図を下に載せます。ついでにすでに掲載した他のホスホリパーゼの切断部位も付け加えておきます。
一般的なリン脂質を加水分解する酵素ホスホリパーゼの種類と、それぞれの酵素が切断する(加水分解する)部位
Gタンパク質のGs、Gi、Gqの3つのGタンパク質を介して何がおきるか、さらにセカンドメッセンジャーは何かをまとめておきましょう。
1)GsはAC活性化を行うことによって、細胞内のcAMPを上昇させます。
2)GiはACを抑制して細胞内のcAMPを減少させます。
3)GqはホスホリパーゼC(PLC)を活性化して、IP3(イノシトールトリフォスフェイト)とDG(ディアシログリセロール)を増やします。
IP3(inositol 1,4,5-trisphosphate訳してイノシトール1,4,5-トリスリン酸)とDAG(Diacyl-glycerol訳してディアシルグリセロール、略してDG)は何でしょうか?例のごとく、難しいですが着いて来て下さい。
まずすべて細胞の膜はリン脂質から成り立っています。リン脂質(Phospholipid)は、構造中にリン酸エステル部位をもつ脂質であります。両親媒性(水にも油にも溶ける)を持ち、脂質二重層を形成して糖脂質やコレステロールと共に細胞膜の主要な構成成分となるほかに大切な仕事は、生体内でのシグナル伝達にも関わっています。
一般的なリン脂質は、グリセリンやスフィンゴシンを中心骨格として脂肪酸とリン酸が結合し、さらにリン酸にアルコールがエステル結合した構造をもっています。アルコールには通常何らかの形で窒素が含まれます。脂肪酸やアルコールには様々な分子種があるため、組み合わせによってきわめて多くの種類が存在します。リン酸は3価の酸であるため、3つのヒドロキシル基のうち2箇所が骨格ならびにアルコールとエステル結合を形成しても、残り1個所は電離してアニオン(陰イオン)が生じます。構造中に疎水性の脂肪酸エステル部位と親水性のリン酸アニオン部位が共存するために、リン脂質は界面活性剤(石鹼)のような両親媒性を示し、水中では外側に親水性部を向けて疎水性部同士が集まることでベシクル(リボソーム)状の安定な脂質二重層を形成します。
リン脂質は、大きく分けてグリセリンを骨格とするグリセロリン脂質と、スフィンゴシンを骨格とするスフィンゴリン脂質の2つが存在します。グリセリンのC1、C2位に脂肪酸がエステル結合し、かつC3位にリン酸がエステル結合した分子をホスファチジン酸となり、ホスファチジン酸からC2位の脂肪酸が外れた分子をリゾホスファチジン酸というのはすでに述べたことがあります。C1には飽和脂肪酸が、C2位には不飽和脂肪酸が結合している場合が多く、アルコールの種類としてはコリンやエタノールアミンやイノシトールやセリンやグリセリンなどがあり、すべてOH基(ヒドロキシル基)をもっています。
スフィンゴシンはパルミチン酸とセリンから合成される物質で、グリセリンのC2位のヒドロキシ基がアミノ基で置き換わり、さらにC1位に長鎖アルキル基が結合した構造を持っています。このため、C2位は脂肪酸とアミド結合(カルボニル基のCがNと結合)を形成します。スフィンゴリン脂質としてはスフィンゴミエリンが知られます。
グリセロリン脂質の生合成は、まずアルコールがキナーゼとアデノシン三リン酸 (ATP) によってリン酸エステル化されます。次にシチジン二リン酸 (CTP) と反応し、活性アルコールとなります。これが1,2-ジグリセリドと反応することによって、グリセロリン脂質が生成します。次にホスファチジルセリンはホスファチジルエタノールアミンのメチル化によっても生じます。
スフィンゴリン脂質(スフィンゴミエリン)は、以前は スフィンゴシンのアミノ基がアセチルCoAによってアセチル化されてセラミドが生じ、次にヒドロキシル基がシチジン二リン酸 (CTP) によって活性化されたコリンと反応してスフィンゴミエリンが生成します。
リン脂質の役割は自己組織化によって自然に脂質二重層を形成し、細胞膜の主要な構成要素になる他に、細胞膜内外の物質移動に用いられる小さな脂質ベシクル(リポソーム)を形成します。脂質二重層は浸透性があり、柔軟で、流体のような特性をもつため、脂質二重層の中にあるリン脂質やタンパク質は面内方向に比較的自由に動くことができます。 また、リン脂質がホスホリパーゼA2などの酵素によって分解されて生じるホスファチジン酸やリゾホスファチジン酸、あるいはアラキドン酸などの各種脂肪酸は、シグナル伝達において重要な役割を担っています。
主なリン脂質には、上に挙げた3つ以外に4つを列挙しておきます。他にもいくつかあります。
1)レシチンと言われるホスファチジルコリン(Phosphatidylcholine、略してPC) があり、アセチルコリン神経系でのアセチルコリン生合成経路におけるコリンの供給源となっています。
2)ホスファチジルエタノールアミン(Phosphatidylethanolamine、略してPE)があり、セファリン(ケファリン、Cephalin)ともいいます。
3)ホスファチジルイノシトール(Phosphatidylinositol、略してPtdIns、またはPI) は細胞膜中の存在量は多くないのですが、PI3キナーゼなどの基質となり、シグナル伝達におけるセカンドメッセンジャーとして働きます。ちなみにPtdInsは、Phospha-tidyl-inositolの先頭のPhospha のPと真ん中のtidyl のtdと、最後のinositol のinsを抜き出して略語にしたものです。
4)ホスファチジルセリン(Phosphatidylserine、略してPS)は、ホスファチジルエタノールアミンをカルボキシル化した構造を持ち、エタノールアミンをセリンに変えただけです。
5)ホスファチジルグリセロール(Phosphatidylglycerol、略してPG、プロスタグランジンと間違えないように)は、植物の葉などに多く含まれます。肺の界面活性剤および原形質膜に見られるグリセロリン脂質です。
6)ジホスファチジルグリセロール(Diphosphatidyl glycerol)は、5)のホスファチジルグリセロールが2つ結合したものです。カルジオリピン(Cardiolipin略してCL)ともいいます。
カルジオリピンとは何でしょうか?
ミトコンドリアの内膜や植物の葉緑体のチラコイド膜にも含まれます。エネルギー通貨であるATPを産生する電子伝達鎖を構成するミトコンドリア内膜の重要な成分がカルジオリピンであり、カルジオリピン(ジホスファチジルグリセロール)は、人体の総脂質組成の約20%を占めています。カルジオリピンはほとんどの細菌の膜でも見つけることができます。カルジオリピンのカルジオという接頭語は、1940年代初頭に牛の心臓から分離されたからです。カルジオリピンはさまざまなミトコンドリアタンパク質の活性発現に必要であり、ミトコンドリア機能の発現に不可欠のリン脂質であります。またコンタクトサイトなどのミトコンドリア特有の膜構造を構築するなど、ミトコンドリアに絶対不可欠の固有のリン脂質であり、リン脂質が2分子重合した特徴的な構造を持っています。これがジホスファチジルグリセロール(Diphosphatidyl glycerol)という名前の由来です。
コンタクトサイト(Contact Site、略してCS)というのは、日本語で接触部位と訳しますが、生体膜間のリン脂質輸送において、二つの生体膜を直接結びつけるタンパク質複合体によって作られる膜間の接触部位が能動的に特別に作られ、ほぼ水に不溶な脂質が小胞輸送を介さずに、小胞体(ER)膜やミトコンドリア内膜等で生合成された後、脂質が溶けない水溶液区画を通過して、各生体膜の特異的脂質組成を確保しつつ細胞全体にくまなく行きわたるために、能動的にMAM(mitochondria-associated ER membrane、日本語でミトコンドリア関連小胞体膜)という膜間コンタクトサイト(CS)を形成する因子と機構が存在していることも解明されました。カルジオリピンの代謝異常は、ミトコンドリア機能の不全を引き起こし、さまざまな疾病の原因となります。
7)スフィンゴミエリン(Sphingomyelin、略してSPH)は髄鞘に多く含まれます。スフィンゴミエリンは、スフィンゴ脂質の一種であり、動物の細胞膜中に存在しており、特に神経細胞の軸索を膜状に覆うミエリン鞘の構成成分として皆さんご存知でしょう。ヒトにおいては、体内に存在するスフィンゴ脂質全体量のうちの85%近くが神経軸索を取り巻く髄鞘を形成するスフィンゴミエリンであります。
既に膜のリン脂質の一つである、ホスファチジルイノシトール(Phosphatidylinositol、略してPI)について述べました。実はこのホスファチジルイノシトールは極めて大切なセカンドメッセンジャーであるジアシルグリセロール(di-acyl-glycerol、略してDAG、別名ジグリセリド)とイノシトール3リン酸(inositol 1,4,5-trisphosphate、略してIP3)を産生する出発点となります。DAGは、グリセリンに2つの脂肪酸がエステル結合を介して結合した分子であります。IP3は、イノシトールに3つのリン酸がエステル結合した化学物質であります。ジアシルグリセロールと共に、細胞のシグナル伝達においてセカンドメッセンジャーの1つとして生体に利用されています。また、どちらも脂質メディエーターとしても使われます。
下に出発点のホスファチジルイノシトール(Phosphatidylinositol、略してPI)が、PI キナーゼ(PI kinase)という酵素によってATPがADPになる時に生じたリン酸をPIにくっつけて、2番目のPI phosphate略してPIPになります。さらに、PIPキナーゼ(PIP kinase)という酵素によってATPがADPになる時に生じたリン酸をPIPにくっつけて、3番、目のPI bis-phosphate略してPIP2になります。さらに反応が進み、DAGとIP3の2つのセカンドメッセンジャーといわれる化学物質ができますが、この反応にはキナーゼが使われるのではないのです。キナーゼはリン酸を付加する反応を触媒します。下の絵図を見ればわかるように、PIP2からDAGとIP3を産生するのに用いられる酵素は、ホスフォリパーゼC-βで、英語でphospho-lipase C-βです。ホスホリパーゼC(phospho-lipase C略してPLC)は、リン酸エステル基の直前でリン脂質を切断する酵素群の総称です。人間を含む真核生物の細胞生理学で、とりわけシグナル伝達経路において重要な役割を果たしています。13種類の哺乳類ホスホリパーゼCは構造に従って、6種類のアイソタイプ(β, γ, δ, ε, ζ, η)に分類されますが、DAGとIP3を産生する時には、6種類のアイソタイプのうちのβが用いられるので、ホスフォリパーゼC-βとなるのです。
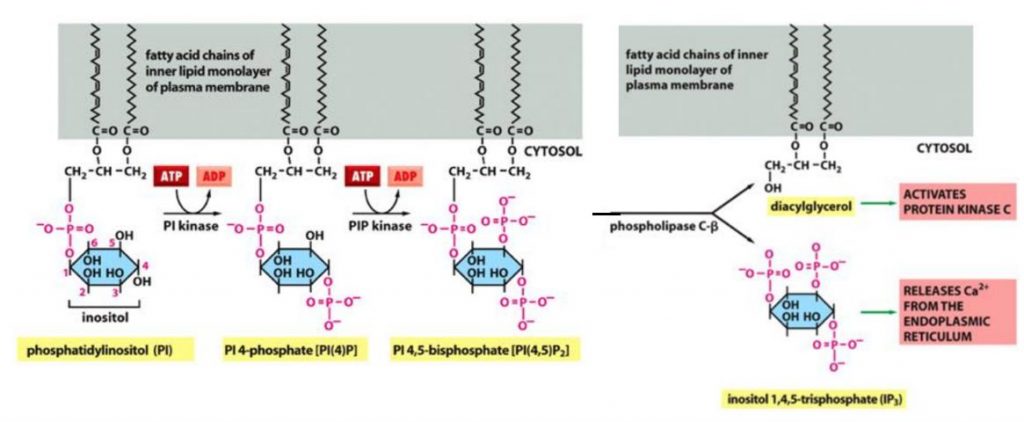
さらに、下に2回も掲載した一般的なリン脂質を加水分解する酵素ホスホリパーゼの種類と、それぞれの酵素が切断する(加水分解する)部位について3回目の掲載をしておきましょう。ホスホは、リン酸という意味であり、リパーゼは脂肪(加水)分解酵素という意味です。脂肪と脂質は同じと考えてください。ついでにイノシトールの構造式も掲載しておきましょう。
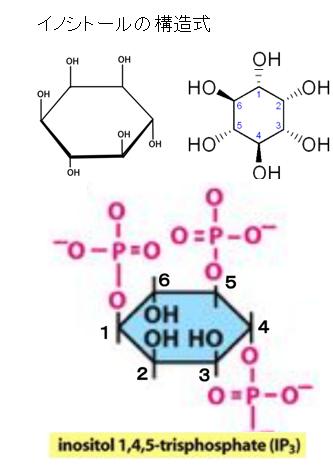
イノシトールとは何でしょうか?
構造式の表記はいくつかあります。もともと構造式は透視図法のような立体を平面に射影した図表ではなく、トポロジー(位相幾何学)的な情報以外は図示されません。すなわち個々の原子と連結した原子との相対的な位置関係以外は図表に表現されないのです。したがって、種々の立体表示方法と投影法が考案されていることを知っておいてください。構造式の左には六員環の炭素には番号がつけられていませんが、右には番号がつけられています。番号の意味を説明しましょう。IP3の正式な名前は、イノシトール3リン酸、inositol 1,4,5-trisphosphateでしたね。Tris-phosphateの意味は、triが3つであり、phosphateがリン酸です。3つのリン酸が1番目と4番目と5番目の炭素に結合しているという意味です。左にinositol 1,4,5-trisphosphateの番号入り構造式を示しておきます。
そもそもイノシトール (inositol) は、シクロヘキサン(六員環)の各炭素上の水素原子が1つずつヒドロキシ基に置き換わった構造の1,2,3,4,5,6-シクロヘキサンヘキサオールを持っています。ビタミンB群の1種であり、ヒトの場合、糖尿病などが原因で体内でイノシトールが不足すると、神経症状が起こるなどの悪影響が出ます。
